Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
In recent years, with the breakthrough progress of CAR-T cell therapy in the treatment of B-cell malignancies, improving efficacy while overcoming challenges such as antigen escape and T-cell exhaustion has become a central focus in the field. CD22, as a B-cell-specific surface molecule, has long been considered an ideal target for immunotherapy. Its restricted expression on mature B cells and most B-cell lymphomas makes it highly promising for both antibody-based therapies and CAR-T cell therapy. Multiple studies published in 2025 have further expanded the strategies for CD22-targeted therapy. On one hand, the CD19xCD22 bicistronic CAR-T construct, through differential signal integration, enhances anti-tumor activity while reducing T-cell exhaustion, demonstrating potential advantages over traditional single-target designs. On the other hand, a domestically pioneered "sandwich" regimen (CD22/CD19 CAR-T combined with autologous stem cell transplantation) has achieved a 2-year overall survival rate of up to 97% in patients with newly diagnosed B-ALL, marking another significant leap for CD22-targeted therapy from the laboratory to the clinic. This article will start from the structure, signaling pathways, and role of CD22 in disease, and systematically review the theoretical basis and application prospects of CD22 as a therapeutic target in light of the latest clinical advances.
CD22, also known as Siglec-2, is mainly expressed in mature B cells, and is a cell surface adhesion molecule that regulates B cell activation, which helps B cells to regulate the sensitivity of antigen response [1][2][3]. Tedder TF, et al. further demonstrated that CD22 expression is specific in B cells and is regulated by development in both mice and humans [4].
As shown in Figure 1, CD22 is expressed in pro-B cells and pre-B cells, and gradually transferred to the cell surface, but the expression level is lower. The vast majority of human I IgM+ and IgD+ B cells express cell surface CD22. But eventually it’s down-regulated in CD27+ memory B cells, especially plasma cells [5]. In lymphoid tissues, hCD22 is expressed by follicular mantle and marginal zone (MZ) B cells, but only weakly by germinal center B cells. In addition, recent studies have found that CD22 expression is also observed in mouse intestinal acid granulocytes, a novel expression pattern indicating that CD22 may have eosinophil regulation [6].
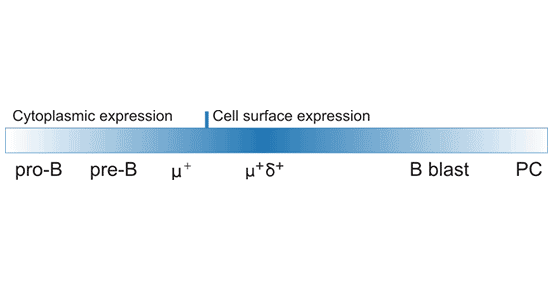
Figure 1. CD22 protein expression during B-cell development and activation.
As shown in Figure 2, the human CD22 gene is located on the long arm of chromosome 19 (19q13.1) with at least 15 exons, of which exon 4-10 encodes a single-chain Ig domain, and exon 11-15 encodes a transmembrane domains and intracellular domains.
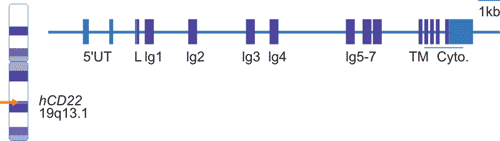
Figure 2. CD22 chromosomal location and organization.
The CD22 protein is a type I transmembrane protein with a molecular weight of 140 kDa and is an important member of the sialic acid-binding immunoglobulin-like lectin family. The extracellular domain of CD22 contains seven Ig domains (Figure 3), and the most distal V-set Ig domain plays a major role in binding to α2,6 sialic acid (α2,6sia) ligands. The function of C2-set Ig domain is to fold the V-set Ig field correctly [7] [8].
The intracellular domain of CD22 includes the immune-receptor tyrosine inhibition motif (ITIM) and the immune-receptor tyrosine activation motif (ITAM) [9]. Ig-like domains 1 and 2 comprise a ligand binding region; when one or more of the six conserved tyrosine residues are phosphorylated, various effector molecules are recruited to the cytoplasmic domain. CD22α and CD22β are two subtypes of CD22, and their extracellular domains have 5 and 7 Ig domains, respectively. These two cDNA subtypes are derived from different splicing of the same gene.
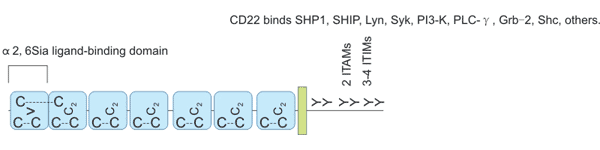
Figure 3. CD22 structure.
CD22 can interact with sialic acid-containing cells, including T cells, B cells, neutrophils, monocytes, and red blood cells [10] [11]. The B cell surface sialic acid ligand which interacts with the CD22 extracellular ligand binding domain is a cis-ligand, and the other cell surface ligands are a trans-ligand. CD22 binds to ligands on the surface of B cells by cis-acting and binds to other cell surface ligands, soluble glycoproteins or cell-linked antigens by trans-acting [12].
The cis-ligand is the major binding ligand for CD22 and is also an important regulator of CD22 activity. As a co-receptor of B cell receptor (BCR), CD22 cross-links CD22 and BCR through antigen, triggers CD22 phosphorylation, dephosphorylating and inactivating downstream signaling proteins. Then BCR signaling is inhibited. CD22 is an essential factor to BCR.
The function of trans-action of CD22 mainly includes Intercellular cross-linking mediation and B cell adhesion and migration regulation. CD22 on the surface of most B cells is obscured, limiting their interaction with the trans-ligand unless the cells are pretreated with sialidase or periodate to bind exogenous sialic acid. Non-masked CD22 may promote its interaction with specific subunits of the trans- or cis-ligand (possibly including BCR).
The ability of CD22 to modulate B-cell signaling is dependent on its proximity to the BCR and is governed by extracellular interactions. As the figure 4 shows, upon B-cell receptor (BCR) cross-linking and translocation to lipid rafts, Lyn phosphorylates the immune-receptor tyrosine-based activation motif tyrosine residues of immunoglobulin a/b [13] [14]. This creates docking sites for other protein tyrosine kinases such as spleen tyrosine kinase (Syk), which in turn phosphorylate and recruit the adaptor molecule BLNK (B-cell linker protein). BLNK forms a scaffold for the association of numerous signaling components, including Vav-1, Btk (Bruton’s tyrosine kinase), and phospholipase Cc2 (PLCγ 2).
Many of these proteins also contain pleckstrin homology domains, and are stabilized in this complex by interaction with the phospholipid PI(3,4,5)P3 (produced by phosphoinositide 3-kinase [PI3K]). Calcium induction upon BCR ligation is brought about by PLCγ 2, which converts PI(4,5)P2 to I(1,4,5)P3 (IP3) and diacylglycerol (DAG). IP3 promotes calcium release from intracellular stores, which in turn triggers an influx of calcium through the opening of Ca2+ release-activated channels (CRACs), and activation of the nuclear factor κB, nuclear factor of activated T cells, and extracellular signal-regulated kinase signaling pathways. Calcium efflux is mediated by the plasma membrane Ca2+ ATPase (PMCA4). [15]
CD22 serves to inhibit the BCR signal at several points in this signaling cascade. Following phosphorylation of its immune-receptor tyrosine-based inhibitory motif residues by Lyn, CD22 provides a docking site for the recruitment of SHP-1 [16]. SHP-1 acts to dephosphorylate and inactivate Vav-1, BLNK, and CD19 (accompanied by a reduction in PI3K activity) to effect a dampening of the BCR signal. SHP-1 also promotes calcium efflux through activation of PMCA4.
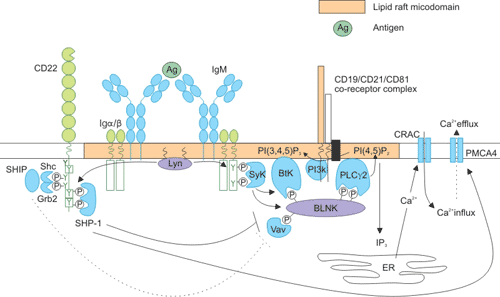
Figure 4. Intracellular signaling pathways of CD22
As mentioned before, The CD22 molecule inhibits the signal produced by the BCR complex recognition antigen [16]. This inhibition was further validated in studies of CD22-deficient mice [17]. B cells in CD22-deficient mice showed mild activation, the number of mature B cells with high IgD and low IgM increased, B cell surface IgM (sIgM) levels decreased, IgM secretion increased and the expression of MHC class II molecules increased [18]. As an antigen that limits B cells, CD22 provides an effective target for the treatment of autoimmune diseases and B cell dysregulation in blood cancer.
CD22 cross-links with BCR, inhibits B cells' response to autoantigens, and prevents autoreactive B cell activation, thereby inhibiting the development of autoimmune diseases. Impaired expression or function of CD22 plays a role in the pathogenesis of autoimmune diseases such as systemic lupus erythematosus (SLE) and rheumatoid arthritis (RA) [19] [20].
SLE is a common chronic systemic autoimmune disease involving multiple organs. Its main features are abnormal activation of polyclonal B cells, autoantibody production, and deposition of immune complexes, causing multiple organ damage. The B cells of CD22-deficient mice are overreacted to the receptor signal, and the increase in Ca2+ influx caused by BCR binding increases the serum IgG anti-dsDNA antibody titer. These antibodies are of polyclonal origin, with poor mutation and high affinity.
In the first part, we have already mentioned that CD22 is only expressed in mature B cells. In most cases, CD22 molecules are still expressed during normal B cell transformation into tumor cells. Related data show that approximately 60-80% of B cell line lymphoma and leukemia cells express CD22 molecules. In addition to chemotherapy, radiation therapy, and naked mAb therapy, radio immunotherapy is currently becoming the fourth treatment for patients with non-Hodgkin's lymphoma. Radiolabeled CD22 mAbs are highly sensitive in the diagnosis and staging of B cell lymphoma. The binding of 131I or 90Y-labeled CD22 mAb (LL2, Eprazumab) to the third Ig domain outside of CD22 completely attenuates antibody- and complement-dependent cytotoxicity mediated by naked CD22 mAb (LL2) in vitro [21] [22].
CD22 molecule is one of the B cell surface inhibitory co-receptors, which is closely related to the development, differentiation and function of B cells. CD22 is restrictedly expressed on the surface of mature B cells and most B lymphoma cells. Taking CD22 as a target, tumor immunotherapy has become one of the hot research topics. Currently, immune-targeting therapy for CD22 molecules, in addition to therapeutic antibodies, also has CAR-T cell therapy. As of November 2019, although only two CD-related antibody drugs have been approved for marketing, there are nearly 108 clinical studies on CD22 CAR-T cell therapy and antibody drugs. The FDA has approved the listing of CD22 related drugs as shown in the following table:
| Drug Name | Mechanism of Action | Drug Type | Therapeutic Indications (Disease Name) | Research Institution/Developer | Highest Development Phase |
|---|---|---|---|---|---|
| Pasotuxizumab | CD22 inhibitor | Antibody-drug Conjugate | Hairy Cell Leukemia | Innate Pharma SA | Approved |
| Inotuzumab Ozogamicin | CD22 inhibitor | DNA inhibitor | ADC | Philadelphia chromosome-positive adult precursor acute lymphoblastic leukemia, etc. | Pfizer Inc. | Pfizer Pharmaceuticals Korea Ltd. | Pfizer Europe MA EEIG | Wyeth Pharmaceuticals LLC | Pfizer (Perth) Pty Ltd. | Approved |
| AUTO-3 | CD19 modulator | CD22 inhibitor | Autologous CAR-T | Tumors | Relapsed/Refractory Central Nervous System Lymphoma | CD19-positive Acute Lymphoblastic Leukemia | Autolus Ltd. | University College London | Phase 2/3 |
| Epratuzumab | CD22 modulator | ADCC | Monoclonal Antibody | Chronic Lymphocytic Leukemia | Lymphoplasmacytic Lymphoma | Multiple Myeloma | Waldenstr枚m's Macroglobulinemia | Immunomedics, Inc. | Phase 2 |
| Recombinant Human CD22 mAb (Sinomab Bioscience / Lonn Ryonn Pharma) | CD22 inhibitor | Monoclonal Antibody | Non-Hodgkin Lymphoma | Systemic Lupus Erythematosus | Alzheimer's Disease, etc. | Sinomab Bioscience (Haikou) Co., Ltd. | China Antibody Pharmaceutical Co., Ltd. | Phase 2 |
| Anti CD22 CAR-T cell therapy (YaKe Biotech) | CD22 inhibitor | CAR-T | Refractory B-cell Acute Lymphoblastic Leukemia | Acute Lymphoblastic Leukemia, etc. | Shanghai YaKe Biotechnology Co., Ltd. | Beijing Gaobo Boran Hospital Co., Ltd. | Phase 2 |
| CD19CD22 CAR-T Therapy (The First Affiliated Hospital of Soochow University) | CD19 modulator | CD22 modulator | Immunocytotoxicity | T lymphocyte substitute | CAR-T | CD19-positive B-cell Acute Lymphoblastic Leukemia, etc. | The First Affiliated Hospital of Soochow University | Phase 2 |
| CD19 and CD22 CAR-T (Yake Biotechnology) | CD19 modulator | CD22 modulator | Immunocytotoxicity | T lymphocyte substitute | Autologous CAR-T | Philadelphia chromosome-positive Acute Lymphoblastic Leukemia | Precursor B-cell Lymphoblastic Leukemia/Lymphoma, etc. | Shanghai YaKe Biotechnology Co., Ltd. | Phase 2 |
| tanCART-19/22 cells | CD19 inhibitor | CD22 inhibitor | CAR-T | Burkitt-like Lymphoma with 11q aberration | CD19-positive Diffuse Large B-cell Lymphoma, etc. | Chinese PLA General Hospital | Phase 2 |
● CD22 protein
Recombinant Human B-cell receptor CD22(CD22),partial (Active) (Code: CSB-MP004900HU)
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
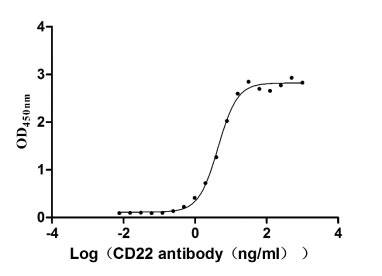
Immobilized CD22 at 2 μg/ml can bind Anti-CD22 rabbit monoclonal antibody, the EC50 of human CD22 protein is 4.034-4.800 ng/ml.
References
[1] Pezzutto A, Rabinovitch PS, et al. Role of the CD22 human B cell antigen in B cell triggering by anti-immunoglobulin [J]. J Immunol. 1988, 140:1791–1795.
[2] Wilson GL, Fox CH, et al. cDNA cloning of the B cellmembrane protein CD22: amediator of B-B cell interactions [J]. J ExpMed. 1991, 173:137–146.
[3] Thomas Dorner, Anthony Shock, et al. CD22 and Autoimmune Disease [J]. International Reviews of Immunology. 2012, 31:363–378.
[4] Tedder TF, Poe JC, et al. CD22: a multifunctional receptor that regulates B lymphocyte survival and signal transduction [J]. Adv Immunol. 2005, 88:1-50.
[5] Daridon C, Blassfeld D, et al. Epratuzumab targeting of CD22 affects adhesion molecule expression andmigration of B-cells in systemic lupus erythematosus [J]. Arthritis ResTher. 2010, 12:R204.
[6] Wen T, Mingler MK, et al. The pan-B cell marker CD22 is expressed on gastrointestinal eosinophils and negatively regulates tissue eosinophilia [J]. J Immunol. 2012, 188:1075–1082.
[7] Engel P, Wagner N, et al. Identification of the ligand-binding domains of CD22, a member of the immunoglobulin superfamily that uniquely binds a sialic acid-dependent ligand [J]. J ExpMed. 1995, 181:1581–1586.
[8] Nath D, van derMerwe PA, et al. Theamino-terminal immunoglobulin like domain of sialoadhesin contains the sialic acid binding site: comparison with CD22 [J]. J Biol Chem. 1995, 270:26184–26191.
[9] Wilson GL, Fox CH, et al. cDNA cloning of the B cell membrane protein CD22: a mediator of B-B cell interactions [J]. J ExpMed. 1991, 173:137–146.
[10] Engel P, Nojima Y, et al. The same epitope on cD22 of B lymphocytes mediates the adhesion of erythmcytes, T and B lymphocytes, neutrophils, and monocytes [J]. J Immunol. 1993, 150(11):4719-4732.
[11] Ramya TN, Weerapana E, et al. In situ trans ligands of CD22 identi6ed by glycan-protein photocross-linking-enabled proteomics [J].Mol Cell Pmteomics. 2010, 9(6):1339-1351.
[12] Smith KGC, Tarlinton DM, et al. Inhibition of the B cell by CD22: a requirement for Lyn [J]. J ExpMed. 1998, 187:807-811.
[13] Xu Y, Harder KW, et al. Lyn tyrosine kinase: accentuating the positive and the negative. Immunity [J]. 2005, 22:9–18.
[14] Doody GM, Justement LB, et al. A role in B cell activation for CD22 and the protein tyrosine phosphatase SHP [J]. Science. 1995, 269:242–244.
[15] Gerlach J, Ghosh S, et al. B cell defects in SLP65/BLNK-deficient mice can be partially corrected by the absence of CD22, an inhibitory coreceptor for BCR signaling [J]. Eur J Immunol. 2003, 33:3418–3426.
[16] June Ereño-Orbea, Taylor Sicard, et al. Molecular basis of human CD22 function and therapeutic targeting [J]. Nature Communications. 2017, 8(764).
[17]Samardzie T, Marinkovic D, et al.Reduction of marginal zone B cells in CD22-deficient mice [J]. Eur J Immunol. 2002, 32(2):561-567.
[18] Tooze RM, Doody GM, et al. Counterregulation by the coreceptom CDl9 and CD22 of MAP kinase activation by membrane immunoglobulin [J]. Immunity. 1997, 7(1):59-67.
[19] O Keefe TL, Williams GT, et al. Hyperresponsive B cells in CD22-de6cient mice [J]. Science. 1996, 274(5288):798-801.
[20] Nakiri Y, Minowa K, et al. E1pression of CD22 on peripheral B cells in patients with rheumatoid arthritis: relation to CD5-positive B cells [J]. Clin Rheumatol. 2007, 26(10):1721-1723.
[21] Thomas F. Tedder, Jonathan C. Poe, et al. CD22: A Multifunctional Receptor That Regulates B Lymphocyte Survival and Signal Transduction [J]. Advances in immunology. 2005, 88:1-50.
[22] Carnahan, J., Wang, P., et al. Epratuzumab, a humanized monoclonal antibody targeting CD22: Characterization of in vitro properties [J]. Clin. Cancer Res. 2003, 9:3982S–3990S.


Comments
Leave a Comment