Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
ATP-binding cassette (ABC) transporters are a family of transmembrane proteins found in all living organisms, from bacteria to humans. These transporters are involved in the translocation of a wide range of substrates across biological membranes [1][2]. They utilize energy derived from ATP hydrolysis to actively transport substrates against their concentration gradient. They are also associated with various diseases, such as cystic fibrosis, Tangier disease, and several forms of cancer.
The substrate-binding domains of ABC transporters are highly variable and determine the specificity of the transporter. There are two main types of ABC transporters: importers, which transport substrates into the cell, and exporters, which transport substrates out of the cell. Importers typically have a extracellular substrate-binding domain that binds the substrate and delivers it to the transmembrane domain for transport. Exporters, on the other hand, have a cytoplasmic substrate-binding domain that binds the substrate and delivers it to the transmembrane domain for export. The importer-type ABC transporters are limited to prokaryotes, while exporter-type ABC transporters are widely found in all organisms.
ABC transporters have a typical modular structure, consisting of four protein domains or subunits: two hydrophobic transmembrane domains (TMDs) and two hydrophilic cytoplasmic nucleotide-binding domains (NBDs) (also called cytoplasmic ABCs).
The TMDs form a channel through the membrane, and the NBDs bind and hydrolyze ATP to provide energy for substrate transport. The mechanism of transport varies depending on the type of ABC transporter, but generally, the substrate binds to the TMDs, ATP binds to the NBDs, and ATP hydrolysis causes a conformational change that drives the substrate across the membrane.
The TMDs form transmembrane translocation pathways or channels through which substrates cross the membrane. ABC importers usually contain 12 transmembrane helices and ABC exporters have 10 transmembrane helices in each TMD. ABC importers need a substrate-binding protein (SBP) that delivers the hydrophilic substrates into the TMD-forming transport pathway. The TMDs control the transported substrates' specificity and direction (influx or efflux).
The NBDs consist of RecA-like sub-domain and helical sub-domain. The P-loops (Walker-A motifs) located in the RecA-like sub-domain and the LSGGQ motif presented in the helical sub-domain are involved in ATP binding and hydrolysis [3]. The NBDs bind and hydrolyze ATP to fuel the transport cycle [2].
The NBDs are conserved throughout the family and possess several featured motifs including Walker A and B motifs that are common to many nucleotide-binding proteins and the ABC signature that is unique to the family, like stacking aromatic D, H, and Q loops.
Many ABC transporters also include extra accessory domains in addition to the four core domains. These comprise cytoplasmic regulatory and catalytic domains as well as extracytoplasmic or membrane-embedded domains.
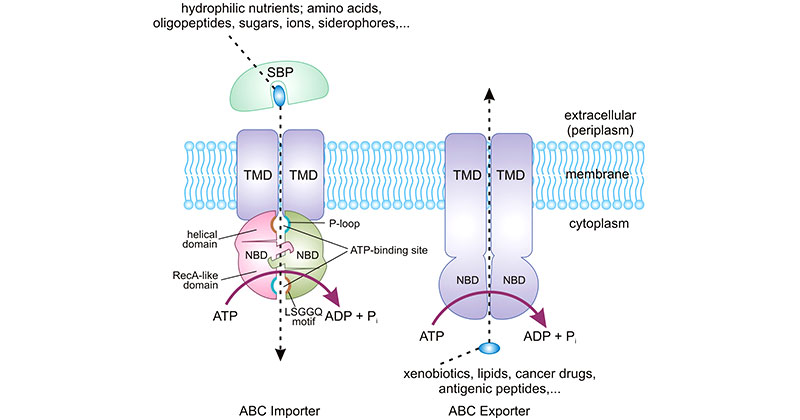
Figure 1. Schematic diagram of the core structure of ABC transporters
The ABC transporter superfamily transfers a variety of substrates, including sugars, amino acids, metal ions, peptides, and proteins, as well as a large number of hydrophobic compounds and metabolites across the extracellular and intracellular membranes.
In bacteria and archaea, there is a periplasmic protein, ABC transporter substrate-binding protein (SBP), that assists in transporting substrates. SBPs bind to specific substrates and delivers them to ABC transporters for transport across the cell membrane. SBPs are involved in the uptake of nutrients such as sugars, amino acids, and metals. They are highly specific for their substrates and can bind to them with high affinity and specificity [16].
ABC transporters are one of the largest known protein superfamilies: there are 49 ABC transporters in humans and 80 in gram-negative E. coli bacteria.
The ABC gene in the human genome is divided into 7 subfamilies (ABCA~ABCG) based on amino acid sequence similarity and phylogeny. The following table is a list of human ABC genes.
Table 1. List of human ABC genes and function
| Subfamily | Member | Function |
|---|---|---|
| ABCA | ABCA1 | Cholesterol efflux onto HDL |
| ABCA2 | Drug resistance | |
| ABCA3 | Phosphatidyl choline efflux | |
| ABCA4 | N-retinylidiene-PE efflux | |
| ABCA5; ABCA6; ABCA7;ABCA8; ABCA9; ABCA10; ABCA11; ABCA12; ABCA13. | / | |
| ABCB | ABCB1 | Multidrug resistance |
| ABCB3 | Peptide transport | |
| ABCB4 | PC transport | |
| ABCB5 | Iron transport | |
| ABCB6 | Fe/S cluster transport | |
| ABCB11 | Bile salt transport | |
| ABCB2; ABCB7; ABCB8; ABCB9; ABCB10. | / | |
| ABCC | ABCC1 | Drug resistance |
| ABCC2 | Organic anion efflux | |
| ABCC3 | Drug resistance | |
| ABCC4 | Nucleoside transport | |
| ABCC5 | Nucleoside transport | |
| CFTR (ABCC7) | Chloride ion channel | |
| ABCC8 | Sulfonylurea receptor | |
| ABCC9 | Potassium channel regulation | |
| ABCC6; ABCC10; ABCC11; ABCC12. | / | |
| ABCD | ABCD1 | VLCFA transport regulation |
| ABCD2; ABCD3; ABCD4. | / | |
| ABCE | ABCE1 | Elongation factor complex |
| ABCF | ABCF1; ABCF2; ABCF3. | / |
| ABCG | ABCG1 | Cholesterol transport |
| ABCG2 | Toxin efflux, drug resistance | |
| ABCG4 | Cholesterol transport | |
| ABCG5 | Sterol transport | |
| ABCG8 | Sterol transport |
ABC transporters import nutrients necessary for cell metabolism and survival, export drugs outside cells, and eliminate toxic compounds inside cells. They also play an essential role in various cell functions, including gene regulation, lipid metabolism modulation, surface motility, cell division and morphology, interspecies competition, outer membrane formation and function, capsule assembly, and chemotaxis.
They are also important in the body's core function of resisting foreign bodies and detoxification [4]. ABC transporters secrete cytotoxic molecules from cells imparting resistance to antibiotics, herbicides, and chemotherapeutic drugs.
Several atypical ABC transporters exert special roles. For example, SMC1-6 is involved in chromosome maintenance. Rad50 participates in DNA and telomere repair. Elf1p takes part in mRNA trafficking.
In eukaryotes, ABC genes are expressed as exporters or efflux transporter proteins responsible for the output of compounds (often toxic molecules or drugs) from the cytoplasm to the extracellular or extracellular compartment (endoplasmic reticulum, mitochondria, peroxisomes). The drug efflux effect of ABC transporters protects the cell from toxins.
Apart from drug efflux, ABC transporters are shown to play a role in drug metabolism. ABC transporters are involved in Phase 0 and Phase III of drug disposition. In phase 0, ABC transporters will control the entry and exit of drugs before they reach their intracellular targets, leading to an increase or decrease in the pharmacological effects of the drug. In phase III, ABC transporters function to ensure the complete elimination of metabolized molecules.
However, some ABC transporters do not directly participate in moving substrates. In the cystic fibrosis transmembrane regulator (CFTR) and the sulfonylurea receptor (SUR), ATP hydrolysis is likely to be associated with the modulation of the opening and closing of the ABC transporter- or other protein-carrying ion channels.
In prokaryotes, ABC genes are expressed either as importers or influx proteins that are implicated in the transport of cell-requiring metabolites such as sugars and amino acids or basic components for biomolecule synthesis such as sulfate and phosphate, or efflux proteins that extrude toxins and drugs from the cells.
In bacteria, the role of ABC transporters is mainly in pathogenicity and toxicity. Bacteria employ these functions to evade or resist the host's defense. Therefore, many cell surfaces or secreted factors can be useful targets for antibacterial therapy or vaccine development.
In plants, ABC transporters are important membrane proteins used to transport a variety of compounds, including heavy metals, antibiotics, phytohormones, and secondary metabolites [5]. The number of ABC transporter family members in the plant genome has more than doubled compared to animals and insects.
In plants, ABC transporters participate in a multitude of physiological processes such as cell detoxification, cuticle formation, stomatal regulation, seed germination, and resistance to pathogenic bacteria, thus enabling the plants to adapt to the adapt to changing environments and cope with biotic and abiotic stresses.
In rice, several ABC transporters of the ABCG family are essential for cuticle formation [6].
In tomatoes, the ABC transporter is involved in the transport of tomato fruit auxin. As a member of the subfamily ABCB, SlABCB4 plays an important role in the transport of auxin during tomato fruit development.
In Arabidopsis, some members of the ABC transporter ABCB subfamily also are involved in auxin transport [7].
ABC transporters also play an important role in the detoxification of pesticides.
Previous study has shown that the ABC transporter acts an important detoxifying enzyme in Plutella xylostella [8].
Epis et al. [9] demonstrated the role of ABC transporters in insecticide defense.
The ABC transporters transport different substrates on the cell membrane. The genetic variation of the ABC transporter superfamily genes is the cause or contributing factor of various Mendelian diseases and complex genetic diseases in humans. The heterozygous variation in ABC transporter gene mutations is related to the susceptibility to specific complex diseases.
Currently, there are 17 ABC genes related to Mendelian inheritance. These include adrenoleukodystrophy, cystic fibrosis, retinal degeneration, hypercholesterolemia and cholestasis, neurological diseases, anemia, and drug reactions. Many of the ABC genes that cause disease lead to many different clinical phenotypes. For example, cystic fibrosis associated with specific CFTR genotypes includes pancreatic sufficiency and pancreatic insufficiency.
One of the functions of ABC transporters is to modulate lipid metabolism. As aberrant lipid metabolism is related to the pathogenesis of metabolic diseases, ABC transporters take part in the suppression of metabolic diseases, including atherosclerosis (AS), coronary artery disease (CAD), hypoalphalipoproteinemia (HA), and Tangier disease (TD).
ABC transporters also have impacts on inducing multidrug resistance (MDR), especially in cancer settings in which ABC transporters can increase the efficacy of anti-cancer drugs.
ABCA1 mediates the transport of intracellular free cholesterol and phospholipids through the cell membrane to poor apoA1 to form HDL [10]. It plays a key role in reverse cholesterol transport and cellular lipid efflux is the initial rate-limiting step of reverted cholesterol transport (RCT) in macrophages [11] and is the most critical determinant of plasma high-density lipoprotein (HDL) levels in hepatic cells.
Mutations in the ABCA1 gene may induce Tangier disease [12] and familial hypolipoproteinemia [13] and may also result in loss of cellular cholesterol homeostasis in prostate cancer. In the general population, some common single-nucleotide polymorphisms of ABCA1 also affect blood lipid levels, atherosclerosis generation, and the severity of coronary heart disease [14].
In recent years, a large amount of evidence indicates that abnormal cholesterol metabolism may play an important role in the development of Alzheimer's disease (AD) (ABCA2/ABCA7), and the apoE gene is also closely related to the metabolism of cholesterol in the brain [15].
Mutations in the ABCA4 gene lead to retinitis pigmentosa, recessive pyramidal malnutrition, and Stargardt disease [17]. All of these diseases are characterized by retinal degeneration, the severity of which is roughly related to the predicted severity of ABCA4 mutation.
Patients with a heterozygous mutation of the ABCA4 gene are more likely to suffer from late-onset macular degeneration-age-related macular degeneration (AMD) [18]. Women with heterozygous mutations of liver transporters that lead to cholestasis may have a higher risk of cholestasis during pregnancy [19].
ABCG subfamily members ABCG5 and ABCG8 are involved in the development of glutathione, a genetic disorder of lipid metabolism. ABCG1 mainly mediates the transport of intracellular free cholesterol to mature HDL. It works synergistically with ABCA1 to promote extracellular lipids out of the cell and complete reverse cholesterol transport in the body [20].
ABCG1 deficiency can also participate in foam cell formation, endothelial cell dysfunction, and inflammatory response, and thus affects the formation and development of atherosclerosis.
Table 2. Diseases and phenotyes caused by ABC genes
| Gene | Mendelian disorder | Complex disease | Animal model |
|---|---|---|---|
| ABCA1 | Tangier disease, FHDLD | HDL levels | Mouse, chicken |
| ABCA3 | Surfactant deficiency | / | / |
| ABCA4 | Stargardt/FFM, RP, CRD | AMD | Mouse |
| ABCA12 | Lamellar ichthyosis | / | / |
| ABCB1 | Ivermectin sensitivitya | Digoxin uptake | Mouse, dog |
| ABCB2 | Immune deficiency | / | Mouse |
| ABCB3 | Immune deficiency | / | Mouse |
| ABCB4 | PFIC-3 | ICP | / |
| ABCB7 | XLSA/A | / | / |
| ABCB11 | PFIC-2 | / | / |
| ABCC2 | Dubin-Johnson Syndrome | / | Rat, sheep, monkey |
| ABCC6 | Pseudoxanthoma elasticum | / | Mouse |
| ABCC7 | Cystic Fibrosis, CBAVD | Pancreatitis, bronchiectasis | Mouse |
| ABCC8 | FPHHI | Mouse | |
| ABCC9 | DCVT | ||
| ABCD1 | ALD | Mouse | |
| ABCG5 | Sitosterolemia | Mouse | |
| ABCG8 | Sitosterolemia | Mouse |
This table derived from literature "Evolution of the ATP-binding cassette (ABC) transporter superfamily in vertebrates" [21]
Although ABC transporters can eliminate some unnecessary or life-threatening molecules, these effects are not always beneficial. In the case of bacteria or cancer cells, ABC transporters are also responsible for resistance to antibiotics or chemotherapy.
ABC transporters can mediate the export of various drugs from cancer cells, which reduces the accumulation of anti-tumor drugs in tumor cells, lowing the effective concentration of intercellular drugs thus leading to drug resistance in tumor cells. Some ABC transporters such as MRP1 can not only excrete drugs directly, but also contribute to intracellular isolation of drugs, making drugs cannot bind to the target, which indirectly leads to drug resistance. These two drug efflux mechanisms confer bacteia or cancer cells drug resistance in combination.
Multidrug resistance (MDR) refers to the phenomenon that tumor cells are cross-resistant to a variety of structurally unrelated chemotherapeutic drugs. The efflux of cytotoxic drugs mediated by ATP-binding cassette transporters is the "classical MDR" pathway [23]. Overexpression of ABC transporters in tumor cells is the main mechanism of tumor MDR. Currently, chemotherapy is one of the approaches to cancer treatment, and MDR is a major obstacle to successful chemotherapy [22].
There are four main types of ABC multidrug transporter: P-glycoprotein (P-gp, MDR1, ABCB1), multidrug resistance-associated protein (MDR-related protein /MRP, ABCC), lung resistance-related protein (LRP), and breast cancer resistance protein (BCRP, ABCG2) [24].
Substantial evidence has demonstrated that ABC transporters ABCB1/MDR1/p-glycoprotein, ABCC1/MRP1, and ABCG2/BCRP, exert essential roles in the MDR of cancer cells.
Acquired MDR of hepatocellular carcinoma may be related to the expression of MRP1, MRP3, and MRP5 genes. MRP2 is the main target of endogenous drug resistance. Liver cancer MDR mediated by ABC transporter severely limits chemotherapy efficacy and prognosis.
Pancreatic cancer is resistant to a variety of chemotherapeutic drugs, and resistance is associated with the ABC transport vector superfamily [26].
Table 3. Relationship and resistance mechanism of three drug-resistance protein and tumor
| ABC Transporters | Involved in Tumor Drug Resistance | Related Pathways | Drug Resistance Mechanism | |
|---|---|---|---|---|
| Similarity | Difference | |||
| ABCB1/MDR1/p-gp | Stomach cancer, colon cancer, breast cancer, ovarian cancer, kidney cancer, uterine cancer, leukemia, lymphatic cancer, oral squamous cell carcinoma, pancreatic cancer | PI3K/AKT, JNK, PKC p38MAPK, MEK/ ERK/RSK Wnt/β-catenin | All of them belong to ATP-dependent ABC transporters, which use ATP to pump& drugs outside the cell, resulting in decreased intracellular drug concentration and drug resistance | Redistribute drugs in cells and accumulate drugs into unrelated organelles to reduce drug concentration at target sites, leading to drug resistance; has an inhibitory effect on apoptosis |
| ABCB1/BCRP | Ovarian cancer, leukemia, esophageal cancer, liver cancer, breast cancer | PTEN/PI3K/AKT | Directly participate in drug resistance by pumping chemotherapeutic drugs out of the cell through its active transport function | |
| MRP | Liver cancer, stomach cancer, lung cancer, kidney cancer, pancreatic cancer, breast cancer, ovarian cancer, cervical cancer, prostate cancer, acute leukemia | Notch, PTEN/ PI3K/AKT | Develop resistance by transporting GSH-X complex and participating in vesicle transport; Redistribute drugs within the cell away from the target, indirectly leading to resistance; Cause resistance by altering membrane permeability or changing intracellular PH |
|
Tumor drug resistance mediated by ABC transporters is an important cause of clinical chemotherapy failure. Therefore, effective inhibition of ABC transporters can reverse tumor multidrug resistance. For MDR induced by ABC transporters, inhibition of ABC transporter-mediated drug efflux is the easiest and most direct route.
Inhibitors of ABC transporters include competitive and non-competitive inhibitors. Currently, there are 3 generations of chemical reversals, most of which are competitive inhibitors of MDR1.
First-generation drugs include verapamil, tamoxifen, cyclosporine A, and quinine. Calcium channel blocker verapamil inhibits the synthesis and activity of MDR1 at the mRNA level while competing for MDR1 binding sites, thereby reversing drug resistance [27]. However, achieving effective serum concentration requires large doses of verapamil, which is prone to toxic effects, thus limiting its clinical application.
Second-generation drugs include biricodar, elacridar, and valspodar. They are characterized by stronger reversal activity, low effective dose, and less susceptibility to toxic reactions. However, they have failed to show overall efficacy improvements in multiple randomized clinical trials due to poor efficacy and increased toxicity.
Third-generation drugs are non-competitive inhibitors that can directly bind to the ABC transporters and make them lose the drug efflux function, thus suppressing the intracellular pumping of anti-cancer drugs out of the cell, increasing the accumulation of intracellular anti-cancer drugs, and reversing the MDR of tumors. They have high transporter affinity and low pharmacokinetics, including Tariquidar, Zosuquidar, and Laniquidar.
Strategies for circumvention of MDR also include small interfering RNA (siRNA) [28] and microRNA (miRNA) to down-regulate the expression of ABC transporters.
Plant anti-tumor drugs provide new ideas for the development of MDR reversal agents due to their small side effects, natural sources, and low cost.
It has been found that artemisinin, quercetin, magnolia officinalis phenol, emodin, Zhejiang fritillaria alkaloid, osmanthus cnidii, ginsenoside, total saponin of Panax notoginseng, the root of mahonia mahogany, Ganoderma lucidum and other traditional Chinese medicines or their effective components can reverse the drug resistance of tumor cells by down-regulating the expression of MDR1.
Psoralen, tetramethylpyrazine, tetrandrine, and paeonol are calcium antagonists, which can inhibit the function of pumping drugs out of cells by binding to P-gp, thus increasing the concentration of intracellular chemotherapeutic drugs and reversing resistance.
Studies have found that diosgenin can reverse the resistance of leukemia cells by inhibiting NF-κB down-regulation of MDR1 [29].
In addition, immunotherapy reversion, gene reversion, somatostatin and its analogs reversion, and Chinese herbal reversion were also used to reverse MDR.
The increased oxidative stress and the activated NF-κB transcription factor may be involved in the inhibition of ABCG1 expression by high glucose.
In conclusion, ABC transporters are found in all domains of life and are essential for many physiological processes, including the uptake of nutrients, the removal of toxic compounds, the efflux of drugs, drug resistance, cholesterol metabolism, and antigen presentation. Their involvement in a wide range of physiological processes makes them important targets for drug development and therapeutic intervention.
References
[1] Momburg F, Roelse J, Howard J C, et al. Selectivity of MHC-encoded peptide transporters from human, mouse and rat [J]. Nature (London), 1994, 367(6464): 648-651.
[2] Higgins C F. ABC transporters: from microorganisms to man [J]. Annu Rev Cell Biol, 1991, 8(1): 67-113.
[3] Hyde S C, Emsley P, Hartshorn M J, et al. Structural model of ATP-binding proteing associated with cystic fibrosis, multidrug resistance and bacterial transport [J]. Nature, 1990, 346(6282): 362-365.
[4] Fletcher J I, Haber M, Henderson M J, et al. ABC transporters in cancer: more than just drug efflux pumps [J]. Nature Reviews Cancer, 2010, 10(2): 147.
[5] Yazaki K, Shitan N, Sugiyama A, et al. Cell and molecular biology of ATP-binding cassette proteins in plants [J]. Int Rev Cell Mol Biol, 2009, 276: 263-299.
[6] Bessire M, Borel S, Fabre G, et al. A Member of the PLEIOTROPIC DRUG RESISTANCE Family of ATP Binding Cassette Transporters Is Required for the Formation of a Functional Cuticle in Arabidopsis [J]. Plant Cell, 2011, 23(5): 1958-1970.
[7] Cho M, Cho H T. The function of ABCB transporters in auxin transport [J]. Plant Signaling & Behavior, 2013, 8(2): e22990.
[8] He W, You M, Vasseur L, et al. Developmental and insecticide-resistant insights from the de novo assembled transcriptome of the diamondback moth, Plutella xylostella [J]. Genomics, 2012, 99(3): 169-177.
[9] Epis S D, Porretta V, Mastrantonio S, et al. Temporal dynamics of the ABC transporter response to insecticide treatment: insights from the malaria vector Anopheles stephensi [J]. Sci Rep, 2014, 4: 7435-7435.
[10] Oram J F, Heinecke J W. ATP-Binding Cassette Transporter A1: A Cell Cholesterol Exporter That Protects Against Cardiovascular Disease [J]. Physiological Reviews, 2005, 85(4): 1343.
[11] Jiang Z, Zhou R, Xu C, et al. Genetic variation of the ATP-binding cassette transporter A1 and susceptibility to coronary heart disease [J]. Molecular Genetics and Metabolism, 2011, 103(1): 0-88.
[12] Santamarina-Fojo S, Remaley A T, Neufeld E B, et al. Regulation and intracellular trafficking of the ABCA1 transporter [J]. Journal of Lipid Research, 2001, 42(9): 1339-1345.
[13] Brookswilson A, Marcil M, Clee S M, et al. Mutations in ABC1 in Tangier disease and familial high-density lipoprotein deficiency [J]. Nature Genetics, 1999, 22(4): 336-345.
[14] Cenarro A, Artieda M, Castillo S, et al. A common variant in the ABCA1 gene is associated with a lower risk for premature coronary heart disease in familial hypercholesterolaemia [J]. Journal of Medical Genetics, 2003, 40(3): 163-168.
[15] Lahiri D. Apolipoprotein e as a target for developing new therapeutics for Alzheimer's disease based on studies from protein, RNA, and regulatory region of the gene [J]. Journal of Molecular Neuroscience, 2004, 23(3): 225-234.
[16] Abbas Maqbool, Richard S.P. Horler, et al. The substrate-binding protein in bacterial ABC transporters: dissecting roles in the evolution of substrate specificity [J]. Biochem Soc Trans 1 October 2015; 43 (5): 1011–1017.
[17] Amalia MartínezMir, Paloma E, Allikmets R, et al. Retinitis pigmentosa caused by a homozygous mutation in the Stargardt disease gene ABCR [J]. Nature Genetics, 1998, 18(1): 11.
[18] Allikmets R. Simple and Complex ABCR: Genetic Predisposition to Retinal Disease [J]. American Journal of Human Genetics, 2000, 67(4): 793-799.
[19] Pauli-Magnus C, Lang T, Meier Y, et al. Sequence analysis of bile salt export pump (ABCB11) and multidrug resistance p-glycoprotein 3 (ABCB4, MDR3) in patients with intrahepatic cholestasis of pregnancy [J]. Pharmacogenetics, 2004, 14(2): 91-102.
[20] Gelissen I C, Harris M, Rye K A, et al. ABCA1 and ABCG1 Synergize to Mediate Cholesterol Export to ApoA-I [J]. Arteriosclerosis Thrombosis & Vascular Biology, 2006, 7(3): 541-541.
[21] Dean M, Annilo T. EVOLUTION OF THE ATP-BINDING CASSETTE (ABC) TRANSPORTER SUPERFAMILY IN VERTEBRATES*[J]. Annu Rev Genomics Hum Genet, 2005, 6(1): 123-142.
[22] Binkhathlan Z, Lavasanifar A. P-glycoprotein Inhibition as a Therapeutic Approach for Overcoming Multidrug Resistance in Cancer: Current Status and Future Perspectives [J]. Curr Cancer Drug Targets, 2013, 13(3):-.
[23] Borst P, Elferink R O. Mammalian ABC transporters in health and disease [J]. Ann Rev Biochem, 2003, 71(1): 537.
[24] Igarashi A, Konno H, Tanaka T, et al. Liposomal photofrin enhances therapeutic efficacy of photodynamic therapy against the human gastric cancer [J]. Toxicology Letters, 2003, 145(2): 133-141.
[26] Nieth C, Priebsch A, Stege A, et al. Modulation of the classical multidrug resistance (MDR) phenotype by RNA interference (RNAi) [J]. Febs Letters, 2003, 545(2): 144-150.
[27] Kohno M, Pouyssegur J. Targeting the ERK signaling pathway in cancer therapy [J]. Annals of Medicine, 2006, 38(3): 200-211.
[28] Abbasi M, Lavasanifar A, Uludag H. Recent attempts at RNAi-mediated P-glycoprotein downregulation for reversal of multidrug resistance in cancer [J]. Medicinal Research Reviews, 2013, 33(1): 33-53.
[29] Wang L, Meng Q, Wang C, et al. Dioscin restores the activity of the anticancer agent adriamycin in multidrug-resistant human leukemia K562/adriamycin cells by down-regulating MDR1 via a mechanism involving NF-κB signaling inhibition [J]. Journal of Natural Products, 2013, 76(5): 909-914.
Aquaporin
Ion Channels
G protein-Coupled Receptor
ATP-binding Cassette
Human Leukocyte Antigen