Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
CD40LG (CD154) is a key co-stimulatory molecule in the TNF superfamily, playing a central role in T cell-dependent B cell activation, antibody class switching, and immune memory formation. This article systematically reviews the molecular mechanisms of the CD40LG-CD40 signaling axis, with a focus on its functions in the NF-κB and JAK-STAT pathways and its regulation by non-coding RNAs. It also summarizes the associations between dysregulation of this signaling axis and diseases such as X-linked hyper-IgM syndrome (XHIGM), autoimmune disorders (e.g., rheumatoid arthritis, systemic lupus erythematosus), acute liver injury, and atherosclerosis, aiming to provide a reference for mechanistic research and precision therapy in CD40LG-related diseases.
2. Mechanism of Action and Signaling Pathways of CD40LG
CD40LG (also known as CD154) is a key co-stimulatory molecule in the immune system and belongs to the tumor necrosis factor (TNF) superfamily. It usually binds to CD40 in trimeric form and plays an important role in T cell-dependent B cell activation, antibody class switching, and immune memory formation [1]. Early studies mainly focused on its expression in activated CD4+ T cells, but subsequent studies have shown that CD40LG can also be found in various cells such as platelets, B cells, endothelial cells, and macrophages, among which platelets are the main source of circulating soluble CD40LG [1,2].
The CD40LG-CD40 axis is not only involved in normal immune responses, but also closely associated with various diseases. CD40LG functional deficiency can cause X-linked hyper-IgM syndrome (XHIGM), leading to antibody class switching disorders and increased susceptibility to infection [3,4]; abnormal activation of this pathway is also involved in autoimmunity, inflammatory responses, atherosclerosis, and tumor immune regulation [1,18,21]. In addition, the role of CD40LG has obvious cell type and disease context dependence, and can exhibit pro-inflammatory, immune-supporting or immunosuppressive effects in different microenvironments [6,7]. Therefore, systematically sorting out the molecular characteristics, signaling mechanisms, disease associations, and drug research progress of CD40LG is of great significance for understanding its biological functions and clinical translational value.
After CD40-CD40LG binding, multiple classic downstream pathways can be activated, among which the most important are NF-κB and JAK-STAT. This signaling axis can be involved in cell proliferation, differentiation, cytokine production, and apoptosis regulation. Studies have shown that CD40 can inhibit hepatitis B virus replication and transcription by activating the JAK-STAT pathway, and further regulate the expression of interferon-stimulated genes such as BST2, suggesting that it plays an important role in antiviral immunity [8]. In addition, NF-κB and apoptosis-related signals are also closely related to CD40LG; in the study of pneumonia treatment mechanisms, CD40LG has been identified as a key target in the NF-κB and apoptosis pathways [9].
In addition to classic protein signaling, miRNAs are also an important level regulating the CD40/CD40LG axis. Studies have found that miR-424 and miR-503 can directly target the 3'UTR of CD40 and inhibit its expression in endothelial cells [10]. Under pro-inflammatory stimulation, these two miRNAs are downregulated, leading to increased CD40 expression, thereby promoting inflammation-related angiogenesis. Furthermore, miR-424 and miR-503 are directly regulated by PPARγ, forming a PPARγ-dependent miR-424/503-CD40 axis [10].
In chronic lymphocytic leukemia, the miR-29 family is also involved in CD40 signaling regulation. Downregulation of miR-29a, miR-29b, and miR-29c can enhance the response of CLL cells to CD40 activation and its downstream NF-κB signaling by directly targeting TRAF4 [11]. Meanwhile, BCR signaling can inhibit miR-29 expression through MYC, thereby enhancing CD40-NF-κB signaling, and this regulatory process can be intervened by BCR inhibitors such as ibrutinib and idelalisib [11].
Long non-coding RNAs are also involved in the regulation of the CD40 axis. For example, lncRNA MSTRG.22719.16 can interact with ocu-miR-326-5p through the ceRNA mechanism, thereby regulating CD40 expression. In local thrombosis induced by PMMA bone cement, activation of the MSTRG.22719.16/ocu-miR-326-5p/CD40 axis can reduce thrombosis, suggesting its potential intervention value [12]. Overall, the CD40-miRNA axis is of great significance in cell fate determination and pathological state regulation [13], but its systematic mechanism still needs further study.
Abnormal CD40LG function is associated with a variety of diseases, including primary immunodeficiency, autoimmune and inflammatory diseases, and tumors. In different diseases, the direction and intensity of action of this molecule are significantly affected by cell origin and microenvironment.
XHIGM is the most typical disease type caused by CD40LG deficiency. Patients often present with recurrent infections, including common bacteria as well as opportunistic infections or viral infections such as Pneumocystis jirovecii, Cryptosporidium, Talaromyces marneffei, cytomegalovirus, and adenovirus [3,14,15]. The respiratory tract and biliary system are commonly involved sites, and some patients may also present with interstitial pneumonia or pulmonary alveolar proteinosis as the initial manifestation [16,17]. In addition, patients may be accompanied by autoimmune and inflammatory complications, increasing prognostic risk [18].
The type of CD40LG gene mutation is closely related to clinical phenotype. Some missense mutations can retain CD40L expression but weaken its binding ability to CD40, forming a relatively mild or atypical phenotype [4]; while truncating mutations usually indicate more severe functional defects [17]. Diagnostically, XHIGM cannot rely solely on immunoglobulin profiles, because some patients may present with atypical serological characteristics. Combining genetic testing, flow cytometry, and CD40 binding function analysis can more accurately identify such patients [4,15].
In rheumatoid arthritis, CD40LG is considered to have potential biomarker value. Studies have shown that the level of CD40LG antibody in the serum of RA patients is elevated, especially in patients with interstitial lung disease or severe bone destruction, suggesting that it may be related to disease activity and complication risk [19]. In systemic lupus erythematosus, abnormal activation of the CD40LG-CD40 axis can promote B cell activation and autoantibody production, which is one of the important pathogenic mechanisms [20]. Blocking strategies targeting this pathway have shown potential in alleviating lupus nephritis and overall autoimmune symptoms in animal models [20].
In acute liver injury, the IFNγ-CD40 axis is closely related to intrahepatic microcirculation disorders. IFNγ can upregulate CD40 expression in endothelial cells, promote its interaction with CD40LG, induce tissue factor expression and aggravate coagulation and liver injury [21]. In atherosclerosis, the role of CD40LG shows cell origin differences: T cell-derived CD40L has a greater impact on plaque formation and stability, while platelet-derived CD40L is more important for thrombosis [1]. In addition, the CD40/CD40LG axis also plays an important regulatory role in HBV clearance, local thrombosis, and allergen tolerance [6,8,12].
The role of CD40LG in tumors is dual. Taking breast cancer as an example, low expression in tumor tissue is associated with more advanced stage and worse prognosis, and is significantly associated with infiltration of various immune cells, suggesting that it may be involved in shaping the tumor immune microenvironment [21]. In pan-cancer analysis, CD40LG has also been identified as a T cell priming marker, and its expression is positively correlated with immunotherapy-related features such as MSI-H, high TMB, and high PD-L1 expression [22].
In specific tumors, CD40LG is also involved in disease progression mechanisms. For example, in T-ALL, BACH2 can affect tumor cell survival and proliferation by inhibiting CD28 and CD40LG expression [23]; in Waldenström macroglobulinemia, the CD40/CD40L axis can support a Treg-mediated immunosuppressive microenvironment [7]; in some studies on T-cell lymphoma and oral squamous cell carcinoma, CD40LG has also been suggested to be involved in tumor maintenance or drug response [24-26].
Drug development surrounding the CD40-CD40LG axis is mainly divided into two categories: one is to inhibit this pathway, used in autoimmune diseases, transplant rejection and other scenarios; the other is to activate this pathway to enhance anti-tumor immunity.
Currently, significant progress has been made in the development of drugs targeting CD40LG. A variety of drug types (including fusion proteins, monoclonal antibodies, oncolytic viruses, small molecule drugs, and therapeutic vaccines) have entered clinical studies for indications such as autoimmune diseases, transplant rejection, and various malignancies, with some candidates having advanced to Phase 3 clinical trials. The table below summarizes the representative pipeline programs.
| Drug Name | Target | Drug Type | Indications | Research Institutions | Highest Development Phase |
|---|---|---|---|---|---|
| Dazodalibep | CD40L | Fusion Protein | Sjögren's Syndrome; Lupus Nephritis; Focal Segmental Glomerulosclerosis | Amgen, Inc.; The Ohio State University Wexner Medical Center; Horizon Therapeutics Ltd. (Ireland) | Phase 3 |
| Dapirolizumab pegol | CD40L | Monoclonal Antibody | Systemic Lupus Erythematosus | Biogen, Inc.; UCB Biopharma SRL; UCB Trading (Shanghai) Co., Ltd.; UCB SA | Phase 3 |
| Frexalimab | CD40L | Monoclonal Antibody | Multiple Sclerosis; Secondary Progressive Multiple Sclerosis; Relapsing-Remitting Multiple Sclerosis; Kidney Transplant Rejection, etc. | Sanofi; Sanofi-Aventis Recherche & Développement SA; ImmuNext, Inc. | Phase 3 |
| Tegoprubart | CD40L | Monoclonal Antibody | IgA Nephropathy; Kidney Transplant Rejection; Amyotrophic Lateral Sclerosis; Heart Transplant Rejection | ALS Therapy Development Foundation, Inc.; Eledon Pharmaceuticals, Inc. | Phase 2 |
| Delolimogene mupadenorepvec | 4-1BBL x CD40L | Oncolytic Virus | Bile Duct Cancer; Pancreatic Ductal Adenocarcinoma | Uppsala University Hospital; Lokon Pharma AB | Phase 2 |
| OPT-101 | CD40L | Small Molecule Drug | Community-Acquired Pneumonia; Sepsis | OP LLC | Phase 2 |
| TriMixDC | CD40L x CD70 x TLR4 | Therapeutic Vaccine; Dendritic Cell Vaccine; Melanoma | Melanoma | - | Phase 2 |
| TNX-1500 | CD40L | Monoclonal Antibody | Immunosuppression; Kidney Transplant Rejection; Autoimmune Diseases; Organ Transplant Rejection | Tonix Pharmaceuticals Holding Corp.; Tonix Pharmaceuticals, Inc. | Phase 2 |
| Bria-OTS | CD40L x CD80 x CD86 x CSF-3R x Type I IFN Receptor x IL-12R x IL-7Rα | Therapeutic Vaccine | Recurrent Breast Cancer; Breast Cancer | BriaCell Therapeutics Corp. | Phase 1/2 |
| Adze1.C (Adze Biotechnology) | CD40L | Oncolytic Virus | Melanoma; Metastatic Melanoma; Bladder Cancer; Glioblastoma; Liver Cancer; Triple-Negative Breast Cancer | Adze Biotechnology, Inc.; Adze Biotechnology Australia Pty Ltd | Phase 1 |
| RP-3 | 4-1BB x CD40L x CTLA4 | Oncolytic Virus | Advanced Malignant Solid Tumors | Replimune, Inc. | Phase 1 |
| MEM-288 | CD40L x IFNβ | Oncolytic Virus | Advanced Cancers; BRAF V600E Positive Melanoma; Brain Metastasis; Cutaneous Squamous Cell Carcinoma, etc. | Memgen, Inc. | Phase 1 |
| Tumor-infiltrating Lymphocytes (H. Lee Moffitt Cancer Center and Research Institute) | CD40L | Tumor-Infiltrating Lymphocytes | Non-Muscle Invasive Bladder Cancer; Bladder Urothelial Carcinoma; Advanced Non-Small Cell Lung Cancer, etc. | H. Lee Moffitt Cancer Center & Research Institute, Inc. | Phase 1 |
| Lu-AG22515 | CD40L x albumin | Bispecific Antibody | Graves' Ophthalmopathy; Autoimmune Diseases | H. Lundbeck A/S; Aprilbio Co. Ltd. | Phase 1 |
| Valo D102 | CD40L x OX40L | Recombinant Vector Vaccine; Therapeutic Vaccine; Oncolytic Virus | Melanoma; Non-Small Cell Lung Cancer; Sarcoma; Colorectal Cancer; Cutaneous Melanoma, etc. | Valo Therapeutics Oy | Phase 1 |
| IBI-355 | CD40L | Monoclonal Antibody | Primary Sjögren's Syndrome; Systemic Lupus Erythematosus | Innovent Biologics (Suzhou) Co., Ltd. | Phase 1 |
(Data as of March 19, 2026, sourced from Synapse)
CD40LG is a key molecule in adaptive immune regulation that plays a central role in B cell activation, antibody class switching, and immune memory formation. Its dysfunction is closely associated with XHIGM, autoimmune diseases, inflammatory diseases, and tumors [3, 18, 21]. Meanwhile, CD40LG has dual potential as both a disease marker and a therapeutic target, and its value in precision medicine is receiving increasing attention. CUSABIO provides CD40LG recombinant proteins, antibodies, and ELISA kits to support your research on related mechanisms and targeted drug development.
● CD40LG Recombinant Proteins
Recombinant Human CD40 ligand (CD40LG), partial (Active); CSB-MP004937HU3
Recombinant Mouse CD40 ligand (Cd40lg), partial (Active); CSB-MP004937MO1


● CD40LG Antibodies
CSB-PA06005A0Rb
CSB-PA004937GA01HU
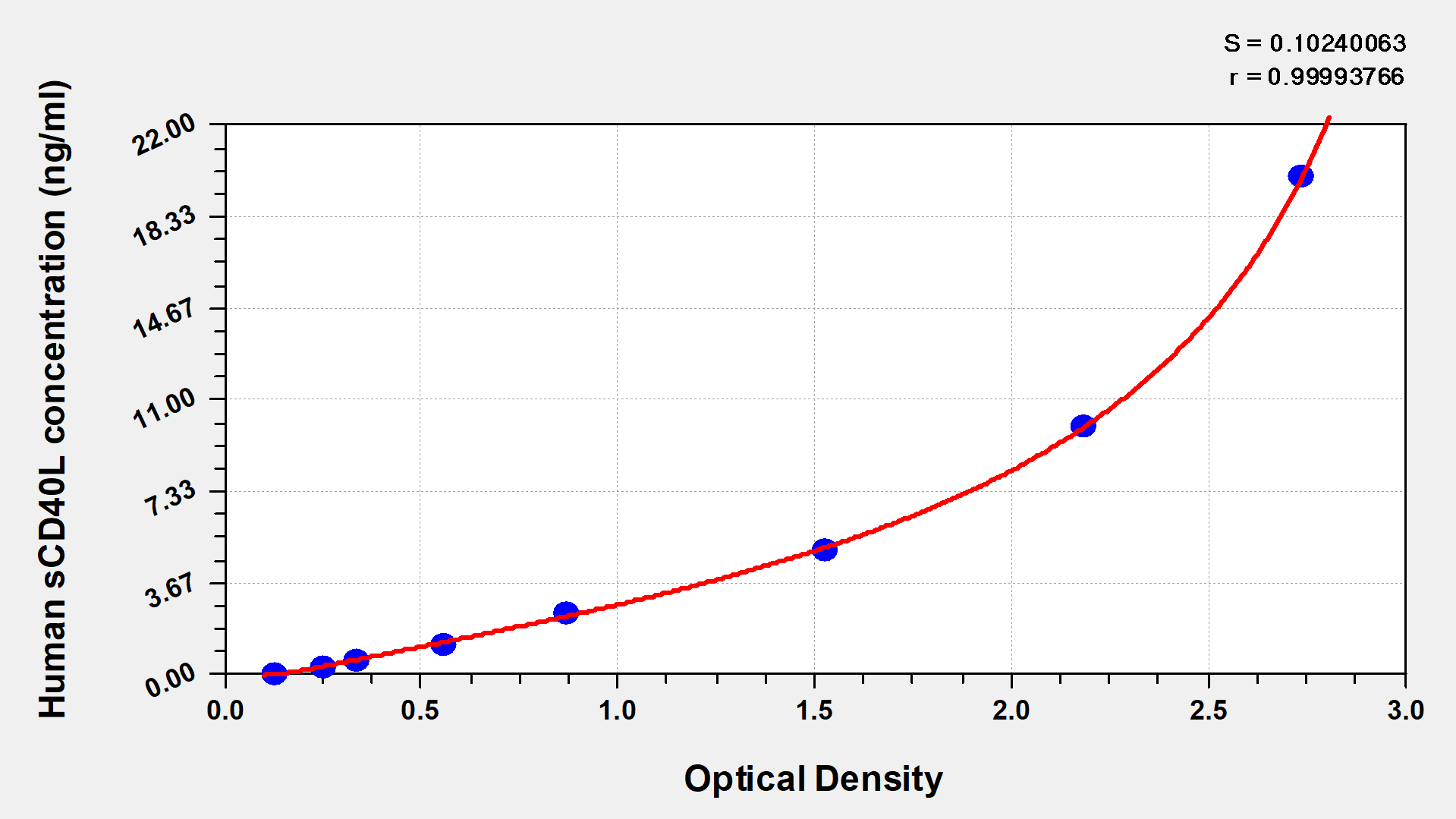
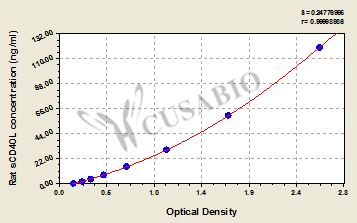
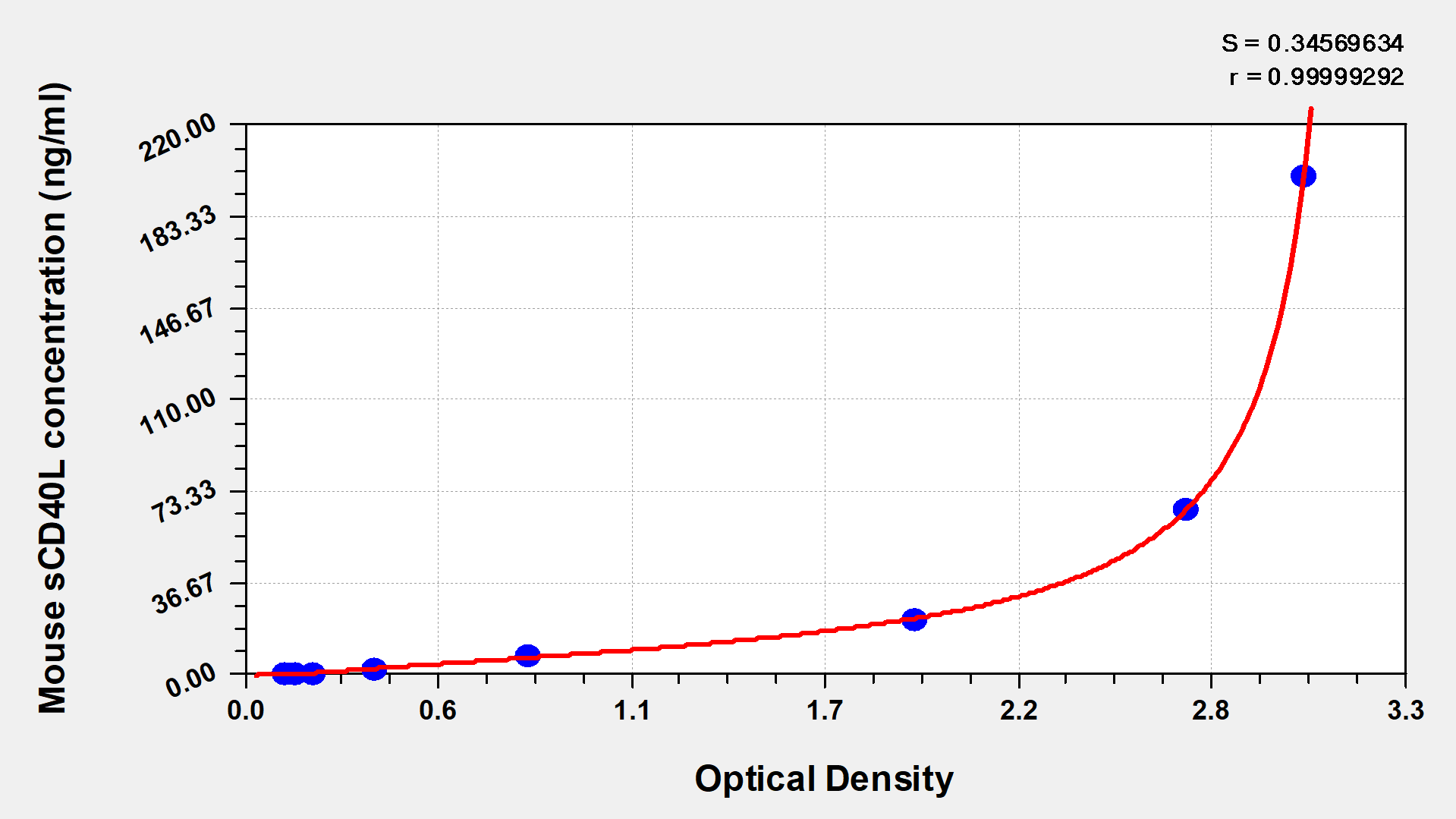
● CD40LG ELISA Kits
References
[1] M. Lacy, C. Bürger, Annelie Shami, M. Ahmadsei, H. Winkels, K. Nitz, C. van Tiel, T. Seijkens, Pascal J. H. Kusters, Ela Karshovka, K. Prange, Yuting Wu, S. Brouns, Sigrid Unterlugauer, Marijke J. E. Kuijpers, M. Reiche, S. Steffens, A. Edsfeldt, R. Megens, J. Heemskerk, I. Gonçalves, C. Weber, N. Gerdes, D. Atzler, E. Lutgens. (2021). Cell-specific and divergent roles of the CD40L-CD40 axis in atherosclerotic vascular disease.
[2] C. Aloui, Antoine Prigent, S. Tariket, C. Sut, Jocelyne Fagan, F. Cognasse, T. Chakroun, O. Garraud, S. Laradi. (2016). Levels of human platelet-derived soluble CD40 ligand depend on haplotypes of CD40LG-CD40-ITGA2.
[3] B. Neven, F. Ferrua. (2020). Hematopoietic Stem Cell Transplantation for Combined Immunodeficiencies, on Behalf of IEWP-EBMT.
[4] T. Nishikawa, Dan Tomomasa, Atsushi Hijikata, Hiroshi Kasabata, Yasuhiro Okamoto, H. Ochs, H. Kanegane. (2025). Case Report: CD40LG Arg203Ile variant underlies atypical phenotype of X-linked hyper IgM syndrome.
[5] K. Antoniou, F. Ender, T. Vollbrandt, Y. Laumonnier, Franziska Rathmann, C. Pasare, Harinder Singh, J. Köhl. (2020). Allergen-Induced C5a/C5aR1 Axis Activation in Pulmonary CD11b+ cDCs Promotes Pulmonary Tolerance through Downregulation of CD40.
[6] A. Sacco, V. Desantis, J. Celay, V. Giustini, Fabio Rigali, Francesco D Savino, M. Cea, D. Soncini, A. Cagnetta, Antonio Giovanni Solimando, D. D'Aliberti, S. Spinelli, Daniele Ramazzotti, C. Almici, K. Todoerti, A. Neri, A. Anastasia, A. Tucci, M. Motta, M. Chiarini, Y. Kawano, Jose-Al Martinez-Climent, R. Piazza, A. Roccaro. (2023). Targeting the immune microenvironment in Waldenström Macroglobulinemia via halting the CD40/CD40-ligand axis.
[7] Jiaxuan Chen, Haitao Chen, Haoming Mai, Shuang Lou, Mengqi Luo, Haisheng Xie, Bin Zhou, J. Hou, Deke Jiang. (2022). A functional variant of CD40 modulates clearance of hepatitis B virus in hepatocytes via regulation of the ANXA2/CD40/BST2 axis.
[8] Liping Sun, Dandan Wang, Yan Xu, Wenxiu Qi, Yanbo Wang. (2020). Evidence of TCM Theory in Treating the Same Disease with Different Methods: Treatment of Pneumonia with Ephedra sinica and Scutellariae Radix as an Example.
[9] . (2017). A PPARγ-dependent miR-424/503-CD40 axis regulates inflammation mediated angiogenesis.
[10] Sonali Sharma, G. Pavlasová, V. Seda, K. Černá, E. Vojáčková, D. Filip, L. Ondrisova, Veronika Šandová, L. Košťálová, P. F. Zeni, M. Borsky, J. Oppelt, K. Lišková, L. Křen, A. Janíková, Š. Pospíšilová, S. Fernandes, M. Shehata, L. Rassenti, U. Jaeger, M. Doubek, M. Davids, Jennifer R. Brown, J. Mayer, T. Kipps, M. Mraz. (2020). miR-29 Modulates CD40 Signaling in Chronic Lymphocytic Leukemia by Targeting TRAF4: an Axis Affected by BCR inhibitors.
[11] Linchao Sang, Luobin Ding, Kangning Hao, Ce Zhang, Xiaoyu Shen, Hui Sun, Dehao Fu, Xiangbei Qi. (2023). LncRNA MSTRG.22719.16 mediates the reduction of enoxaparin sodium high-viscosity bone cement-induced thrombosis by targeting the ocu-miR-326-5p/CD40 axis.
[12] S. Salunkhe, Tushar Vaidya. (2020). CD40-miRNA axis controls prospective cell fate determinants during B cell differentiation.
[13] H. Fan, Li Huang, Diyuan Yang, Changhao Zhang, Q. Zeng, G. Yin, G. Lu, Kunling Shen. (2022). Respiratory infections in X-linked hyper-IgM syndrome with CD40LG mutation: a case series of seven children in China.
[14] A. Khojah, L. Gunderman, Mohammad Binhussein, A. Bukhari, Imad Khojah. (2023). A Case Report of Hyper-IgM Syndrome Patient with Normal Serum IgA Level.
[15] Tianliu Wang, Li-Fang Wu, Junguo Chen, Wen Zhu, Hua Wang, Xiao-Lin Liu, Yi-qun Teng. (2019). X-linked hyper-IgM syndrome complicated with interstitial pneumonia and liver injury: a new mutation locus in the CD40LG gene.
[16] Hong-bo Xu, Maolin Tian, Yong-hua Bai, X. Ran, Lei Li, Yan Chen. (2023). CD40LG-associated X-linked Hyper-IgM Syndrome (XHIGM) with pulmonary alveolar proteinosis: a case report.
[17] W. Rae, D. Ward, C. Mattocks, Yifang Gao, R. Pengelly, Sanjay Patel, S. Ennis, S. Faust, Anthony P. Williams. (2017). Autoimmunity/inflammation in a monogenic primary immunodeficiency cohort.
[18] Chuanhui Yao, Hui Xu, Xun Gong, Xieli Ma, Yuchen Yang, Dan Dou, Qiu-wei Peng, Juan Jiao, Xiaopo Tang, Quan Jiang, Congmin Xia. (2025). CD40LG as a Biomarker in Rheumatoid Arthritis: Links to Bone Destruction and Interstitial Lung Disease ‐ A Bioinformatic Analysis With Clinical Validation.
[19] Tian-Liang Fang, Baoqi Li, Meng Li, Yu-long Zhang, Zhang Jing, Yuan Li, Tianyuan Xue, Zhirang Zhang, Wen-li Fang, Zhongda Lin, Fanqiang Meng, Liyan Li, Yang Yang, Xingding Zhang, Xin Liang, Shu-Na Chen, Jun Chen, Xudong Zhang. (2023). Engineered Cell Membrane Vesicles Expressing CD40 Alleviates System Lupus Nephritis by Intervening B Cell Activation.
[20] M. Kurokawa, T. Goya, M. Kohjima, Masatake Tanaka, Sadahiro Iwabuchi, Shigeyuki Shichino, S. Ueha, Tomonobu Hioki, Tomomi Aoyagi, Masaki Kato, Kouji Matsushima, Yoshihiro Ogawa. (2024). Microcirculatory disturbance in acute liver injury is triggered by IFNγ-CD40 axis.
[21] Manqiu Yuan, Jianying Pei, Ruihao Li, Lirong Tian, Xin-bin He, Yanping Li. (2021). CD40LG as a Prognostic Molecular Marker Regulates Tumor Microenvironment Through Immune Process in Breast Cancer.
[22] H. Miyashita, R. Kurzrock, N. Bevins, Kartheeswaran Thangathurai, Suzanna Lee, S. Pabla, M. Nesline, S. Glenn, J. Conroy, P. DePietro, E. Rubin, J. Sicklick, S. Kato. (2023). T-cell priming transcriptomic markers: implications of immunome heterogeneity for precision immunotherapy.
[23] Min Feng, Bailing Zhang, Guilan Li, Yan Yang, Jiangyuan Liu, Ziting Zhang, Bing Zhou, Han Zhang. (2024). BACH2-mediated CD28 and CD40LG axes contribute to pathogenesis and progression of T-cell lymphoblastic leukemia.
[24] M. Fujisawa, Tran Bich Nguyen, Yoshiaki Abe, Y. Suehara, K. Fukumoto, S. Suma, K. Makishima, C. Kaneko, Yen Thi Minh Nguyen, K. Usuki, K. Narita, K. Matsue, N. Nakamura, S. Ishikawa, F. Miura, Takashi Ito, A. Suzuki, Yutaka Suzuki, S. Mizuno, Satoru Takahashi, S. Chiba, M. Sakata-Yanagimoto. (2022). Clonal germinal center B cells function as a niche for T-cell lymphoma.
[25] Sofia Papadakos, Hawraa Issa, Abdulaziz Alamri, Abdullah Alamri, A. Semlali. (2024). Rapamycin as a Potential Alternative Drug for Squamous Cell Gingiva Carcinoma (Ca9-22): A Focus on Cell Cycle, Apoptosis and Autophagy Genetic Profile.
[26] E. Fukutani, P. I. Ramos, J. I. Kasprzykowski, L. G. Azevedo, M. Rodrigues, J. Lima, Helton Fábio Santos de Araújo Junior, K. Fukutani, A. T. D. de Queiroz. (2019). Meta-Analysis of HTLV-1-Infected Patients Identifies CD40LG and GBP2 as Markers of ATLL and HAM/TSP Clinical Status: Two Genes Beat as One.


Comments
Leave a Comment