Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-E04581m |
| Size | 96T,5×96T,10×96T |
| Price | Request a Quote |
| Trial Size |
24T ELISA Kit Trial Size (Only USD$150/ kit) * Sample kit cost can be deducted as a $30 credit for each 96-assay kit of the same analyte and brand you subsequently purchase within six months until depleted. More details >> Interested in a trial size? Please leave a message below.
|
| Have Questions? | Leave a Message or Start an on-line Chat |
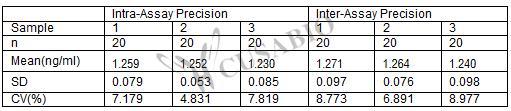
Intra-assay Precision (Precision within an assay): CV%<8%
Three samples of known concentration were tested twenty times on one plate to assess.
Inter-assay Precision (Precision between assays): CV%<10%
Three samples of known concentration were tested in twenty assays to assess.
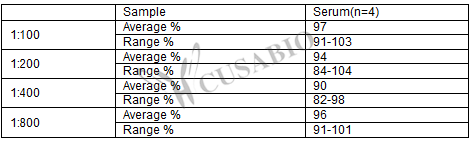
To assess the linearity of the assay, samples were spiked with high concentrations of mouse IGF-1 in various matrices and diluted with the Sample Diluent to produce samples with values within the dynamic range of the assay.
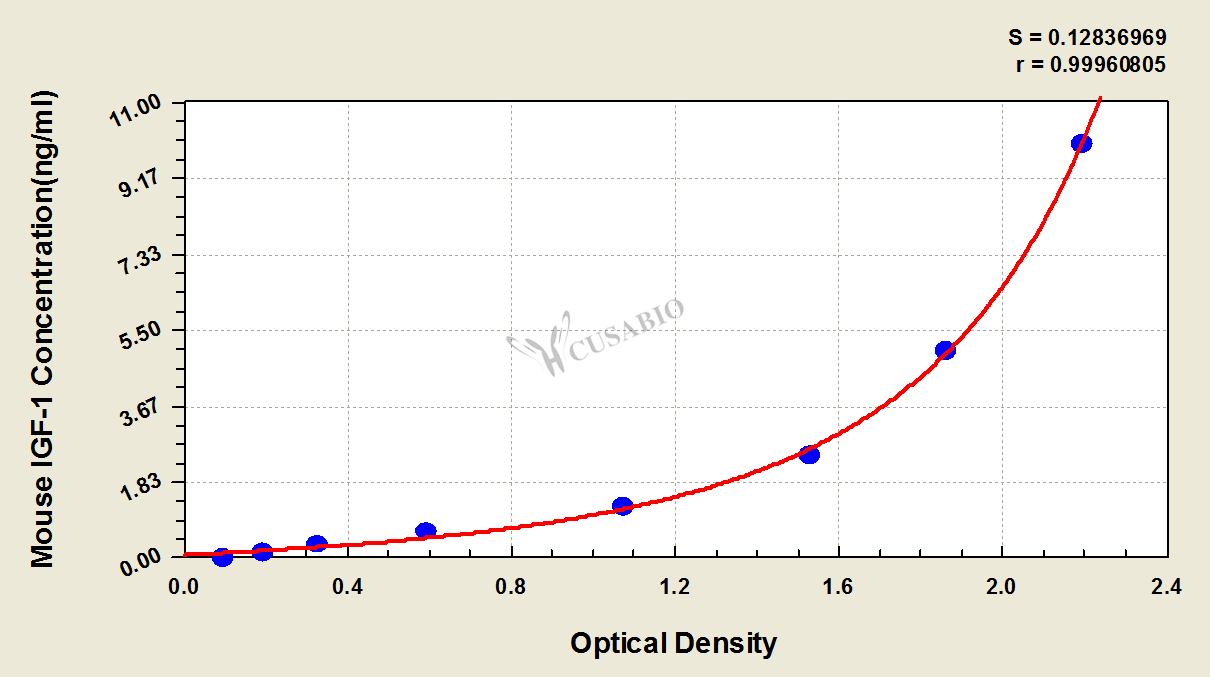
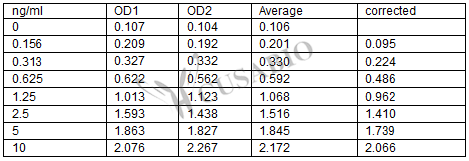
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed.
The mouse IGF1 ELISA Kit is engineered for accurate measurement of mouse IGF1 levels from samples including serum, plasma, or tissue homogenates. It uses the Sandwich-ELISA mechanism in combination with the enzyme-substrate chromogenic reaction to measure the IGF1 content in the sample. The color intensity is positively correlated with IGF1 content in the sample. This kit has been validated against standards of sensitivity, specificity, precision, linearity, recovery, and lot-to-lot consistency.
IGF1 is a polypeptide hormone with endocrine, paracrine, and autocrine effects. IGF-1 gives an inhibitory feedback signal on growth hormone (GH) secretion in the hypothalamus by stimulating somatostatin generation in the pituitary. It is involved in multiple physical actions, including tissue growth & development, proliferation, lipid metabolism, pro-survival/anti-aging, anti-inflammatory, anabolic, antioxidant with neuro- and hepatoprotective properties, as well as the progression of cell cycle and mitogenesis. IGF1 protects mitochondria by preventing them from oxidative damage generated via increased metabolism, enhancing ATP synthesis, and decreasing intramitochondrial production of free radicals. The GH/IGF1/insulin pathway plays an essential role in the pathogenesis of several age-related diseases including cancer, dementia, cardiovascular, and metabolic diseases.
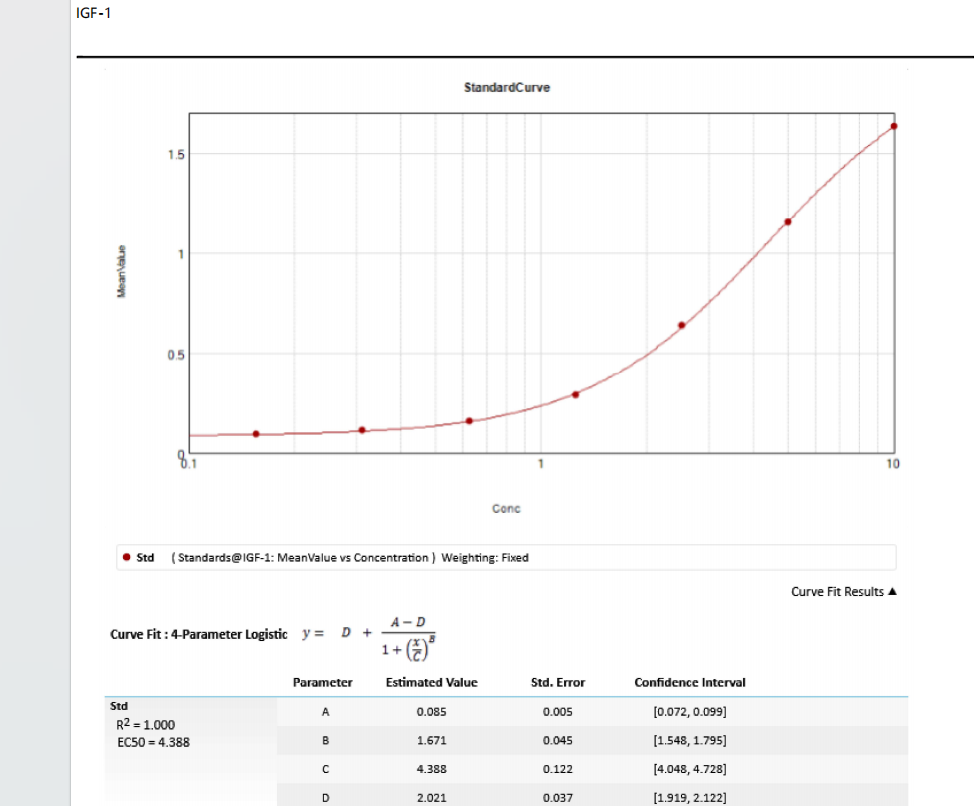
Sample type: Serum
Sample species: Mouse
Sample dilution: 1:100
Review: I used CSB-E04581m to detect mouse igf-1 to detect mouse OD value in the predicted range, the measured curve meets the requirements of the instructions, the experimental operation is simple, the steps are convenient, and the measured results are perfect. It\'s a nice curve.
By Anonymous

