Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
DDIT3, DNA damage-inducible transcript 3, alias CHOP, GADD153, is a transcription factor with a key regulatory role in cellular stress response, especially endoplasmic reticulum stress. It promotes autophagy-related gene expression by interacting with ATF4 and activates cellular autophagy by inhibiting the mTOR signaling pathway.DDIT3 is also involved in apoptosis, inducing mitochondria-associated apoptosis by regulating the Bcl-2 family and C/EBPδ. DDIT3 plays an important role in the regulation of autophagy and apoptosis in tumor cells, and its aberrant expression may increase the sensitivity of tumor cells to therapy and is a potential target for tumor therapy. Under glutamine starvation conditions, DDIT3 balances glycolysis and mitochondrial oxidative phosphorylation through a dual mechanism, reduces reactive oxygen species production, and helps tumor cells to adapt to metabolic stress, demonstrating its biological significance in the metabolic regulation of tumor cells.
DDIT3 Recombinant Monoclonal Antibody (CSB-RA918842A0HU)
Validated Data
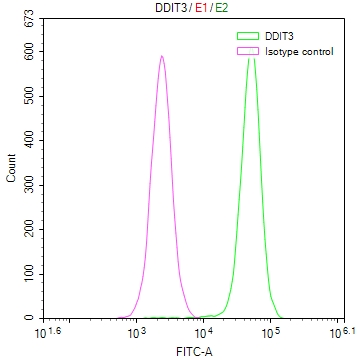
Overlay Peak curve showing Hela cells stained with CSB-RA918842A0HU (red line) at 1:50. The cells were fixed in 4% formaldehyde and permeated by 0.2% TritonX-100. Then 10% normal goat serum to block non-specific protein-protein interactions followed by the antibody (1µg/1*106cells) for 45min at 4℃. The secondary antibody used was FITC-conjugated Goat Anti-rabbit IgG(H+L) at 1:200 dilution for 35min at 4℃.Control antibody (green line) was rabbit IgG (1µg/1*106cells) used under the same conditions. Acquisition of >10,000 events was performed.
The following DDIT3 reagents supplied by CUSABIO are manufactured under a strict quality control system. Multiple applications have been validated and solid technical support is offered.
DDIT3 Antibodies for Homo sapiens (Human)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA006589GA01HU | DDIT3 Antibody | Human,Mouse,Rat | ELISA,WB |
| CSB-PA071811 | DDIT3 Antibody | Human,Mouse,Rat | ELISA, WB, IHC, IF |
| CSB-PA799326 | Phospho-DDIT3 (Ser30) Antibody | Human,Mouse | ELISA,WB |
| CSB-PA001639 | DDIT3 Antibody | Human,Mouse,Rat | WB, IHC, IF, ELISA |
| CSB-PA001640 | DDIT3 Antibody | Human,Mouse | WB, IHC, IF, ELISA |
| CSB-PA007699 | Phospho-DDIT3 (S30) Antibody | Human,Mouse | WB, IHC, IF, ELISA |
| CSB-PA006589LA01HU | DDIT3 Antibody | Human | ELISA, IHC, IF |
| CSB-PA006589LB01HU | DDIT3 Antibody, HRP conjugated | Human | ELISA |
| CSB-RA918842A0HU | DDIT3 Recombinant Monoclonal Antibody | Human | ELISA, FC |
DDIT3 Proteins for Homo sapiens (Human)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006589HU CSB-EP006589HU CSB-BP006589HU CSB-MP006589HU CSB-EP006589HU-B |
Recombinant Human DNA damage-inducible transcript 3 protein (DDIT3) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
DDIT3 Proteins for Mus musculus (Mouse)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006589MO CSB-EP006589MO CSB-BP006589MO CSB-MP006589MO CSB-EP006589MO-B |
Recombinant Mouse DNA damage-inducible transcript 3 protein (Ddit3) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
DDIT3 Proteins for Cricetulus griseus (Chinese hamster) (Cricetulus barabensis griseus)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006589DXU CSB-BP006589DXU CSB-MP006589DXU CSB-EP006589DXU-B |
Recombinant Cricetulus griseus DNA damage-inducible transcript 3 protein (DDIT3) | Yeast Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
DDIT3 Proteins for Bos taurus (Bovine)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP606584BO CSB-EP606584BO CSB-BP606584BO CSB-MP606584BO CSB-EP606584BO-B |
Recombinant Bovine DNA damage-inducible transcript 3 protein (DDIT3) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
DDIT3 Proteins for Rattus norvegicus (Rat)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP737096RA CSB-EP737096RA CSB-BP737096RA CSB-MP737096RA CSB-EP737096RA-B |
Recombinant Rat DNA damage-inducible transcript 3 protein (Ddit3) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |

