Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
E-cadherin, encoded by the CDH1 gene, is a calcium-dependent cell adhesion molecule in epithelial cells, playing a central role in maintaining epithelial structural integrity, intercellular communication, and tissue homeostasis [1,2]. Impaired CDH1 function often leads to the loosening of intercellular adhesion and triggers key phenotypic changes such as epithelial-mesenchymal transition (EMT), making it a significant driver in the development and progression of various tumors [3]. This article provides a structured review of the structure and physiological functions of CDH1, its multi-level regulatory mechanisms, key pathways, associated diseases, and the latest research progress in targeted drugs, aiming to assist in your research endeavors.
1. Background and Research Significance: Why CDH1 is a "Adhesion-Signaling-Disease" Hub
2. CDH1/E-cadherin Structure and Cell-Cell Adhesion: From Molecular Assembly to Mechanical Coupling
4. Key Signaling Pathways and Cellular Phenotypes: From EMT to Microenvironmental Adaptation
E-cadherin mechanically couples adjacent cells by forming adherens junctions and connects to the actin cytoskeleton, participating in cell migration, differentiation, and signal transduction [2]. Therefore, E-cadherin is generally regarded as an important tumor suppressor, and its loss can weaken cell adhesion and promote invasion and metastasis [3]. Beyond macroscopic tissue structure, E-cadherin is also involved in finer spatial organization: in keratinocytes, E-cadherin loss can cause small-scale cell demixing, suggesting that adhesion plays a "fine-tuning" role in microscopic tissue homogeneity/pattern formation [4].
Clinically, pathogenic germline mutations in CDH1 are highly associated with hereditary diffuse gastric cancer (HDGC), with carriers having a significantly elevated lifetime risk of diffuse gastric cancer [5-9]. In specific populations (e.g., New Zealand Māori), it has been confirmed as a major genetic factor contributing to the high incidence of early-onset diffuse gastric cancer [5]. Besides coding region mutations, deletions of downstream regulatory sequences (e.g., CDH1-TANGO6 deletion) can also significantly downregulate CDH1 expression, leading to very early-onset, highly penetrant diffuse gastric cancer [10]. In sporadic gastric cancer, both CDH1 genetic variations and epigenetic alterations (such as promoter methylation) are closely related to the carcinogenic process [11]. Furthermore, "escape" from cell-cell/cell-matrix adhesion dependence in diffuse gastric cancer is associated with RHO signaling perturbations (e.g., RHOA mutations or ARHGAP fusions), emphasizing a synergistic relationship between adhesion changes and intracellular signaling remodeling [12].
Notably, CDH1 may exhibit differential roles in different tumor contexts. For example, in breast cancer, CDH1 expression can be upregulated and is associated with stage, metastasis, stem cell characteristics, and poor prognosis, suggesting it may exert pro-tumorigenic effects in specific contexts [13]. Conversely, in triple-negative breast cancer, where treatment options are limited, CDH1 deficiency is common, prompting the exploration of targeting strategies for "CDH1-deficient tumors" [14].
E-cadherin is a transmembrane glycoprotein. Its extracellular domain consists of multiple cadherin repeats, which stabilize in the presence of calcium ions and mediate homotypic binding, forming the basis of adherens junctions [1]. Its intracellular domain binds to adaptor proteins like β-catenin, linking to the actin cytoskeleton, thereby enabling mechanical transmission and tissue-scale mechanical coupling [17,2]. Related biophysical models conceptualize the adhesion complex as a "spring-like structure" capable of transmitting resistance/tension, which can explain multicellular behaviors such as cell polarization, oscillatory dynamics, and supracellular stress fibers. This suggests that the "adhesion-cytoskeleton" is not a static link but a dynamically coupled system [2].
Structural pathogenic variants can directly disrupt adhesive function. For example, the G212E missense mutation significantly affects E-cadherin stability, localization, and adhesive capacity, leading to tissue disorganization and削弱ing anti-invasive properties [19]. Additionally, E-cadherin function is regulated by complex signaling networks: PAK5, for instance, is involved in maintaining cell-cell adhesion integrity, indicating that the adhesion complex is not merely a "structural component" but a functional module continuously regulated by kinase networks [20].
In HDGC and sporadic gastric cancer, CDH1 expression loss or functional impairment can result from coding region mutations, regulatory sequence deletions, or promoter methylation [5,10,11]. In inflammatory contexts, increased CpG methylation at the CDH1 locus has been reported in association with mucosal inflammation, suggesting chronic inflammation may impair the epithelial barrier through epigenetic pathways [24]. Evidence of aberrant CDH1 methylation and expression loss also exists in breast cancer [25].
Various miRNAs/lncRNAs are implicated in explaining CDH1 dynamics in different tumors. For instance, miR-92a-3p can target CDH1/β-catenin and influence Notch-1/Akt signaling in glioma and glioma stem-like cells, participating in tumor phenotype regulation [26]. LncRNA SNHG1 forms a complex with the RNA-binding protein hnRNPL to co-regulate CDH1, thereby promoting prostate cancer growth and metastasis [32].
Glycosylation is an important mechanism affecting E-cadherin activity and cellular behavior. In pancreatic cancer cells, ST3Gal III alters the sialylation pattern of E-cadherin, reducing cell-cell aggregation while enhancing invasion/migration-related signals (such as FAK Tyr397 phosphorylation), indicating a functional link between "glycan-adhesion-migration" [22]. At the interaction level, the MCC protein can interact with E-cadherin and β-catenin, enhancing adhesion in colorectal cancer cells, suggesting the stability of the adhesion complex also relies on cooperation within the tumor suppressor network [21].
The most typical consequence of CDH1 deficiency is the enhancement of EMT-related phenotypes. Research suggests that SPHK1 can induce EMT by promoting the autophagy-lysosomal degradation of CDH1/E-cadherin, pointing to a "metabolic enzyme - autophagy - adhesion degradation" cascade potentially involved in liver cancer progression [31]. On the other hand, CDH1 expression is also linked to metabolic reprogramming: E-cadherin can induce serine synthesis to support breast cancer progression and metastasis, hinting that under specific conditions, it might promote tumor adaptation through metabolic pathways [28]. ZHX2 deficiency enriches hybrid MET cells and affects the EMT/MET balance by regulating E-cadherin expression, emphasizing that CDH1 is not a simple "on/off" switch but may participate in multi-state transitions [33].
At the pathway level, CDH1 is often intertwined with networks such as Wnt/β-catenin, PI3K/AKT/mTOR, MAPK/ERK, and TGF-β/Smad. Previous research in esophageal cancer discussed epigenetic dysregulation of the Wnt/β-catenin and TGF-β-Smad pathways and their impact on prognosis [3]. In colorectal cancer, genetic and epigenetic alterations in WNT pathway components are associated with microsatellite instability stratification [30].
Furthermore, CDH1 abnormalities are linked to epithelial barrier disruption: In a SARS-CoV-2-infected Caco-2 intestinal epithelial model, CDH1/E-cadherin expression and soluble E-cadherin release were affected, discussed as a potential pathophysiological basis for intestinal manifestations [29].
HDGC represents the scenario with the most closed-loop characteristics regarding "genetics - mechanism - clinical management" in CDH1 research. Current studies emphasize that in the New Zealand Māori population, germline CDH1 mutations significantly contribute to the high frequency of early-onset diffuse gastric cancer, highlighting its significant public health importance in specific genetic backgrounds/population structures [5]. Beyond classic coding region mutations, combined loss of CDH1 and its downstream regulatory sequences (CDH1-TANGO6 deletion) can cause a marked decrease in CDH1 expression and is associated with very early-onset, highly penetrant diffuse gastric cancer, suggesting that "regulatory regions outside the coding sequence" can also determine disease burden [10].
In sporadic gastric cancer, CDH1-related genetic variations and epigenetic alterations (e.g., regulatory region changes and promoter methylation) are discussed together as important factors affecting CDH1 expression and the carcinogenic process, indicating that HDGC and sporadic gastric cancer are not entirely separate entities but may represent a continuum with "different intensities along the same mechanistic axis" [11]. Studies using gastric cancer organoid models emphasize that during disease progression, cells can "escape" dependence on cell-cell and cell-matrix adhesion, accompanied by RHO signaling perturbations (such as RHOA mutations or ARHGAP fusions) [12]. Such evidence links "CDH1 deficiency leading to adhesion imbalance" with "intracellular signaling remodeling," providing mechanistic clues for understanding the aggressive biological behavior of diffuse gastric cancer [12].
Breast cancer exhibits greater heterogeneity regarding CDH1. Germline CDH1 mutations are associated with hereditary lobular breast cancer, relevant for discussing hereditary cancer risk spectra and genetic counseling paths [9]. Additionally, familial breast cancer risk has been reported to be associated with CDH1 allele SNPs [27]. In some studies, CDH1 expression may be upregulated and correlate with stage, metastasis, stem cell characteristics, and poor prognosis, suggesting it might exert pro-tumorigenic effects in certain contexts [13]. This implies that in the context of breast cancer, one cannot simply generalize with "high E-cadherin = tumor suppressor, low = tumor promoter"; clinical interpretation often requires integration with tumor subtyping and molecular networks [13]. In scenarios with limited treatment options like triple-negative breast cancer, CDH1 deficiency is more readily incorporated into discussions of "actionable vulnerabilities," prompting exploration of combination inhibition strategies [14].
Evidence in colorectal cancer leans more towards "genetic susceptibility - network homeostasis - tumor immune context." GWAS studies have included CDH1 in the genetic susceptibility map for colorectal cancer, indicating its risk relevance at the population level [15]. MCC protein interaction with E-cadherin/β-catenin enhances adhesion in colorectal cancer cells, emphasizing that CDH1-related phenotypes are not solely determined by a single gene but are also influenced by the partner protein network of the adhesion complex [21]. In inflammatory bowel disease-associated colorectal cancer, IBD-related genes are analyzed for prognostic and tumor immune implications, with CDH1 also included in relevant discussion frameworks [16]. Corroborating this, alterations in WNT pathway components in colorectal cancer and their relationship with microsatellite instability stratification provide a macro-level pathway background for explaining epithelial stability disruption and signaling reprogramming [30].
In esophageal cancer, reduced expression of CDH1 or CTNNB1 is associated with lymph node metastasis and poor prognosis, linking "adhesion complex imbalance" to less favorable clinical outcomes [17]. Concurrently, epigenetic dysregulation of Wnt/β-catenin and TGF-β-Smad pathways is used to explain prognostic differences in esophageal cancer, suggesting that CDH1 alterations often co-occur with broader pathway-level abnormalities [3].
At the precancerous lesion level, altered expression of EMT-related proteins (including E-cadherin) in oral lichen planus suggests that cell junction disruption may be linked to lesion progression, providing evidence for the "early indicative significance of adhesion changes" [18].
CDH1 is relevant not only to tumors but also to epithelial barrier status. In inflammatory contexts, increased CpG methylation at the CDH1 locus is associated with mucosal inflammation, suggesting chronic inflammation may affect epithelial adhesion and barrier homeostasis through epigenetic mechanisms [24]. In a SARS-CoV-2-infected Caco-2 intestinal epithelial model, changes in CDH1/E-cadherin expression and soluble E-cadherin release were observed and discussed as part of the potential pathological basis for intestinal manifestations [29]. Related to detection, elevated soluble E-cadherin fragments in urine are thought to potentially reflect the shedding/cleavage process from epithelial tumor cells, offering clues for non-invasive biomarkers [23].
Current drug development targeting CDH1 is primarily in preclinical and early discovery stages, encompassing small molecule drugs, biologics, exosomes, etc. Main exploratory directions include malignant glioma, breast cancer, and polycystic diseases, involving institutions such as Shenyang Pharmaceutical University, Harvard University, and Sichuan Cancer Hospital.
| Drug Name | Type | Indications | Development Stage | Research Institution |
|---|---|---|---|---|
| AL-GDa62 | Small Molecule Inhibitor | CDH1-Deficient Gastric Cancer | Preclinical | University of Otago, New Zealand |
| Dasatinib | Multi-Kinase Inhibitor | CDH1-Deficient Tumors | Preclinical | Multiple Research Institutions |
| FAK Inhibitor + ROS1 Inhibitor | Combination Therapy | CDH1-Deficient Cancers | Preclinical | Multiple Research Institutions |
| Exosome-Delivered CDH1 mRNA | Gene Therapy | Epithelial Barrier Damage Repair | Exploratory Phase | Domestic Research Institutions (China) |
CUSABIO offers CDH1 recombinant proteins, antibodies, and ELISA kits to support your mechanistic studies and targeted drug development.
● CDH1 Recombinant Proteins
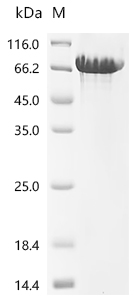
Recombinant Human Cadherin-1(CDH1),partial (Active); CSB-MP005034HU1
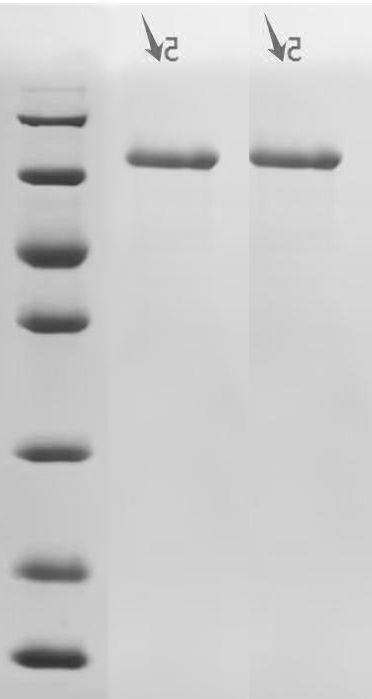
Recombinant Macaca fascicularis Cadherin-1 (CDH1), partial (Active); CSB-MP5601MOV
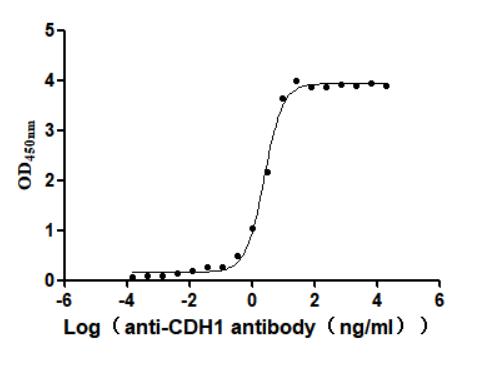
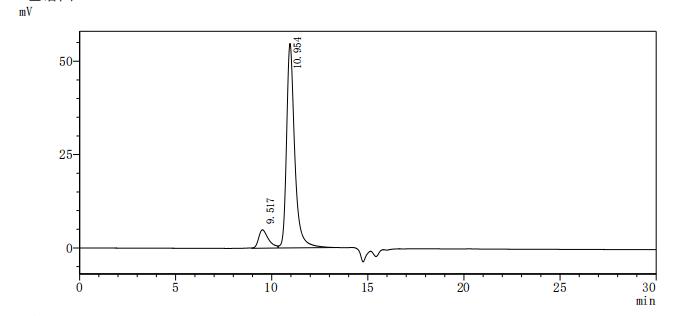
● CDH1 Antibodies
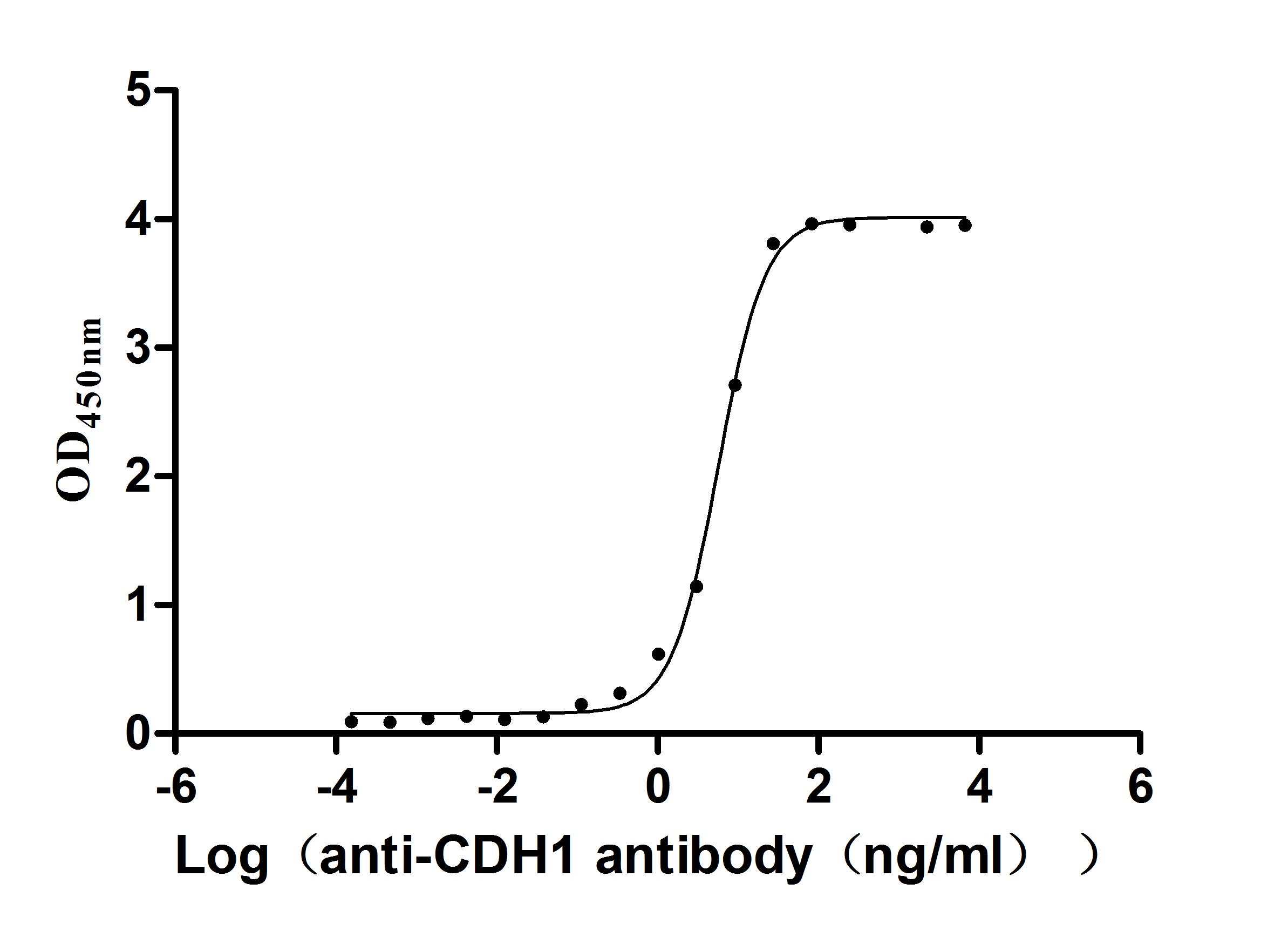
CDH1 Recombinant Monoclonal Antibody; CSB-RA005034MA1HU
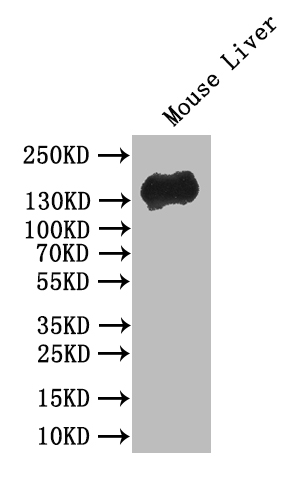
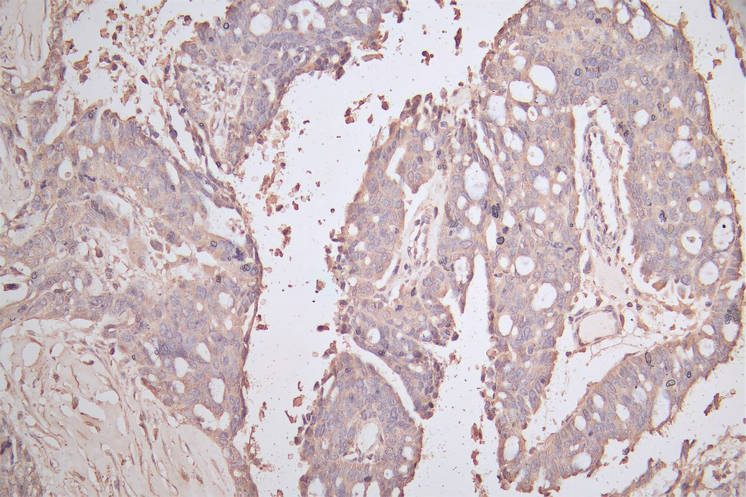
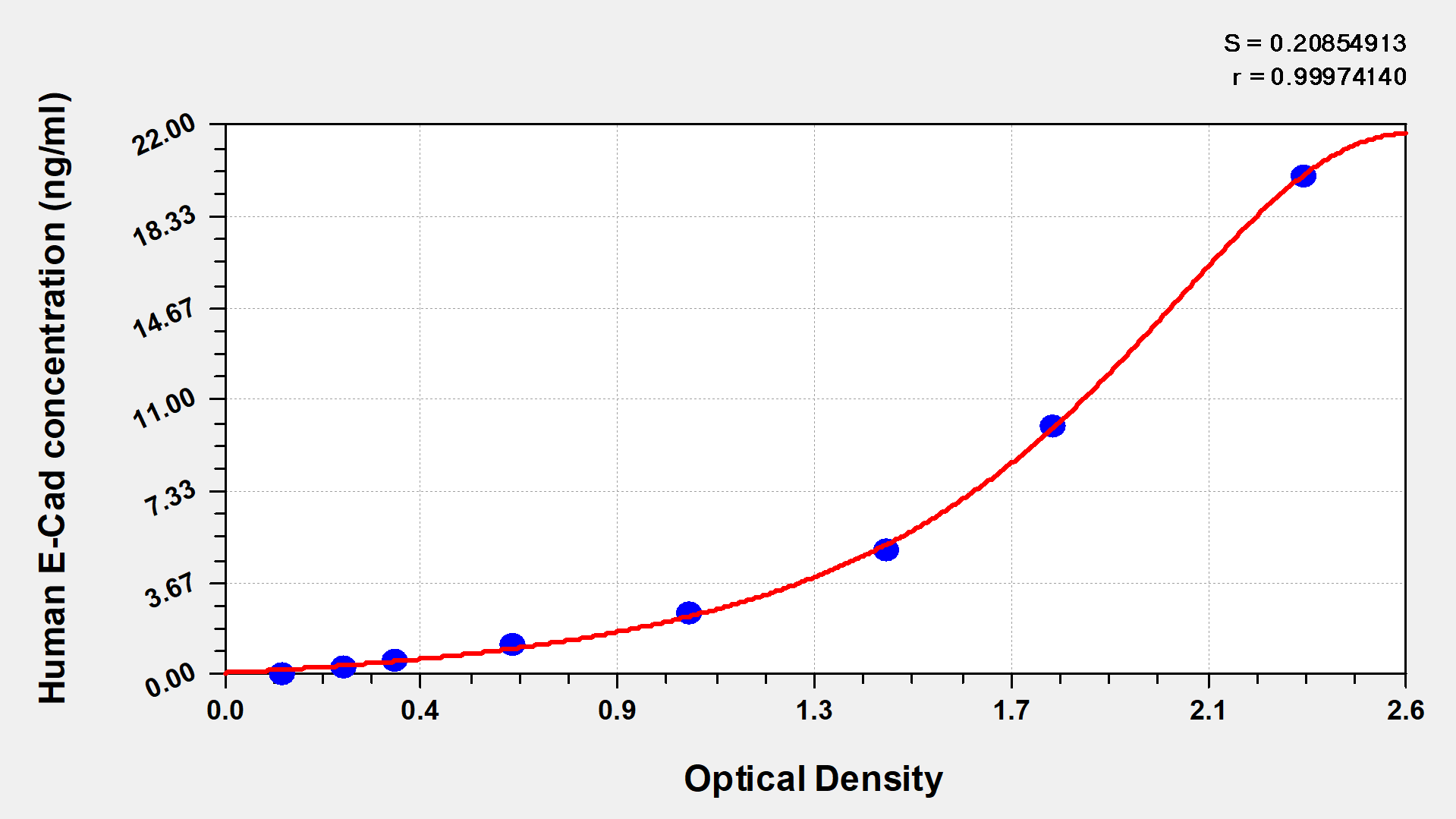
● CDH1 ELISA Kits
References
[1] Anna Zhigun, Mabel Lizzy Rajendran.(2023). Modelling non-local cell-cell adhesion: a multiscale approach.
[2] XinXin Du, Ido Lavi, Michael J. Shelley.(2025). Collective multicellular patterns arising from cadherin-linked cytoskeletal domains.
[3] Virendra Singh, A. Singh, I. Sharma, L. Singh, J. Sharma, B. B. Borthakar, A. Rai, A. Kataki, S. Kapur, S. Saxena.(2019). Epigenetic deregulations of Wnt/β-catenin and transforming growth factor beta-Smad pathways in esophageal cancer: Outcome of DNA methylation.
[4] Preeti Sahu, Daniel M. Sussman, Matthias Rubsam, Aaron F. Mertz, Valerie Horsley, Eric R. Dufresne, Carien M. Niessen, M. Cristina Marchetti, M. Lisa Manning, J. M. Schwarz.(2019). Small-scale demixing in confluent biological tissues.
[5] C. Hakkaart, L. Ellison-Loschmann, R. Day, A. Sporle, Jonathan Koea, Pauline Harawira, Soo Cheng, M. Gray, Tracey Whaanga, N. Pearce, P. Guilford.(2018). Germline CDH1 mutations are a significant contributor to the high frequency of early-onset diffuse gastric cancer cases in New Zealand Māori.
[6] I. Chen, Lesley Mathews-Greiner, Dandan Li, A. Abisoye-Ogunniyan, Satyajit Ray, Yansong Bian, V. Shukla, Xiaohu Zhang, Rajarashi Guha, Craig J. Thomas, B. Gryder, A. Zacharia, J. Beane, S. Ravichandran, M. Ferrer, U. Rudloff.(2017). Transcriptomic profiling and quantitative high-throughput (qHTS) drug screening of CDH1 deficient hereditary diffuse gastric cancer (HDGC) cells identify treatment leads for familial gastric cancer.
[7] Nicola Bougen-Zhukov, Lyvianne Decourtye-Espiard, Wilson Mitchell, Kieran Redpath, J. Perkinson, T. Godwin, M. Black, P. Guilford.(2022). E-Cadherin-Deficient Cells Are Sensitive to the Multikinase Inhibitor Dasatinib.
[8] T. Godwin, S. Kelly, Tom Brew, Nicola Bougen-Zhukov, A. Single, Augustine Chen, Cassie E Stylianou, L. Harris, Sophie K Currie, Bryony J. Telford, Henry Beetham, G. Evans, M. Black, P. Guilford.(2018). E-cadherin-deficient cells have synthetic lethal vulnerabilities in plasma membrane organisation, dynamics and function.
[9] G. Corso, M. Intra, C. Trentin, P. Veronesi, V. Galimberti.(2016). CDH1 germline mutations and hereditary lobular breast cancer.
[10] C. São José, J. Garcia-Pelaez, Marta Ferreira, O. Arrieta, Ana André, Nelson Martins, Samantha Solís, B. Martínez-Benítez, M. Ordoñez-Sánchez, M. Rodríguez-Torres, Anna K. Sommer, Iris B A W Te Paske, C. Caldas, M. Tischkowitz, María Teresa Tusié, Stefan Gabriel Sérgio Richarda M. Gareth Susana José Luzi Aretz Capella Castedo de Voer Evans Fernandes Garc, S. Aretz, G. Capellá, S. Castedo, Richarda M. de Voer, Gareth Evans, Susana Fernandes, J. Garcia-Pelaez, L. Garrido, E. Holinski-Feder, N. Hoogerbrugge, D. Huntsman, A. Jahn, C. Kets, A. Laner, M. Ligtenberg, Andrea Meinhardt, A. Mensenkamp, Carla Oliveira, S. Peters, I. Quintana, E. Schröck, Anna K. Sommer, I. Spier, L. Spruijt, V. Steinke-Lange, I. T. Paske, M. Tischkowitz, L. Valle, R. S. van der Post, Y. V. van Herwaarden, W. V. van Zelst-Stams, D. William, N. Hoogerbrugge, German Demidov, Richarda M. de Voer, S. Laurie, Carla Oliveira.(2023). Combined loss of CDH1 and downstream regulatory sequences drive early-onset diffuse gastric cancer and increase penetrance of hereditary diffuse gastric cancer.
[11] G. Tedaldi, C. Molinari, C. São José, Rita Barbosa-Matos, A. André, R. Danesi, V. Arcangeli, M. Ravegnani, L. Saragoni, P. Morgagni, F. Rebuzzi, M. Canale, S. Pignatta, Elisa Ferracci, G. Martinelli, G. Ranzani, Carla Oliveira, D. Calistri, P. Ulivi.(2021). Genetic and Epigenetic Alterations of CDH1 Regulatory Regions in Hereditary and Sporadic Gastric Cancer.
[12] Yin Tong, P. Cheng, Chung Sze Or, Sarah S K Yue, H. Siu, S. Ho, S. Law, W. Tsui, D. Chan, Stephanie Ma, Siu Po Lee, Annie S. Y. Chan, April S Chan, Shui Wa Yun, Ho Sang Hui, S. Yuen, S. Leung, Helen H. N. Yan.(2022). Escape from cell-cell and cell-matrix adhesion dependence underscores disease progression in gastric cancer organoid models.
[13] D. Xie, Yiyu Chen, Xue Wan, Jingyuan Li, Qin Pei, Yanan Luo, Jinbo Liu, Ting Ye.(2022). The Potential Role of CDH1 as an Oncogene Combined With Related miRNAs and Their Diagnostic Value in Breast Cancer.
[14] Jiaming Gao, Yunying Yao, Zaiqi Wang, Baoyuan Zhang, R. Ren.(2023). Abstract 5821: Synergism of FAK and ROS1 inhibitors in the treatment of CDH1-deficient cancers mediated by FAK-YAP-TRX signaling.
[15] Da-Tian Bau, Ting-Yuan Liu, Jai-Sing Yang, William Tzu-Liang Chen, Chia‐Wen Tsai, Wen‐Shin Chang, Tao-Wei Ke, Chi-Chou Liao, Yu-Chia Chen, Yen-Ting Chang, Fuu-Jen Tsai.(2024). Characterizing Genetic Susceptibility to Colorectal Cancer in Taiwan Through Genome‐Wide Association Study.
[16] Di Wang, Biao Xie.(2022). Prognostic and tumor immunity implication of inflammatory bowel disease-associated genes in colorectal cancer.
[17] H. Ishiguro, T. Wakasugi, Yukio Terashita, N. Sakamoto, Tatsuya Tanaka, K. Mizoguchi, H. Sagawa, T. Okubo, H. Takeyama.(2016). Decreased expression of CDH1 or CTNNB1 affects poor prognosis of patients with esophageal cancer.
[18] L. Hämäläinen, Y. Soini, S. Pasonen-Seppänen, M. Siponen.(2019). Alterations in the expression of EMT-related proteins claudin-1, -4 and -7, E-cadherin, TWIST1 and ZEB1 in oral lichen planus..
[19] J. Figueiredo, F. Mercadillo, Soraia Melo, A. Barroso, Margarida Gonçalves, J. Diaz-Tasende, Patrícia Carneiro, L. Robles, F. Colina, C. Ibarrola, J. Perea, Eurico Morais-de-Sá, R. Seruca, M. Urioste.(2021). Germline CDH1 G212E Missense Variant: Combining Clinical, In Vitro and In Vivo Strategies to Unravel Disease Burden.
[20] A. F. Ismail, Sevil Oskay Halacli, N. Babteen, M. De Piano, T. Martin, W. Jiang, Muhammad S. Khan, P. Dasgupta, C. Wells.(2017). PAK5 mediates cell: cell adhesion integrity via interaction with E-cadherin in bladder cancer cells..
[21] F. Benthani, F. Benthani, D. Herrmann, D. Herrmann, P. N. Tran, P. N. Tran, L. Pangon, L. Pangon, M. Lucas, A. Allam, N. Currey, S. Al‐Sohaily, Marc Giry-Laterrière, J. Warusavitarne, P. Timpson, P. Timpson, M. R. Kohonen-Corish, M. R. Kohonen-Corish, M. R. Kohonen-Corish.(2018). ‘MCC’ protein interacts with E-cadherin and β-catenin strengthening cell–cell adhesion of HCT116 colon cancer cells.
[22] Sònia Bassagañas, S. Carvalho, A. M. Dias, Marta Pérez-Garay, M. R. Ortiz, J. Figueras, C. Reis, S. Pinho, R. Peracaula.(2014). Pancreatic Cancer Cell Glycosylation Regulates Cell Adhesion and Invasion through the Modulation of α2β1 Integrin and E-Cadherin Function.
[23] M. Katayama, S. Hirai, M. Yasumoto, K. Nishikawa, S. Nagata, M. Otsuka, K. Kamihagi, I. Kato.(1994). Soluble fragments of e-cadherin cell-adhesion molecule increase in urinary-excretion of cancer-patients, potentially indicating its shedding from epithelial tumor-cells..
[24] Charles de Ponthaud, Solafah Abdalla, M. Belot, Xiaojian Shao, Christophe Penna, A. Brouquet, Pierre Bougnères.(2024). Increased CpG methylation at the CDH1 locus in inflamed ileal mucosa of patients with Crohn disease.
[25] Asia Asiaf, S. T. Ahmad, S. Aziz, A. Malik, Zubaida Rasool, A. Masood, M. Zargar.(2014). Loss of expression and aberrant methylation of the CDH1 (E-cadherin) gene in breast cancer patients from Kashmir..
[26] Hang Song, Yao-quan Zhang, Na Liu, Sheng Zhao, Y. Kong, Liudi Yuan.(2016). miR-92a-3p Exerts Various Effects in Glioma and Glioma Stem-Like Cells Specifically Targeting CDH1/β-Catenin and Notch-1/Akt Signaling Pathways.
[27] M. Bagherpour, Kamelia Gharibzad, H. Rassi.(2018). Association of CDH1 and TERT Single-Nucleotide Polymorphisms with Susceptibility to Familial Breast Cancer Risk..
[28] Geonhui Lee, Claudia Wong, Anna Cho, Junior J West, Ashleigh J. Crawford, Gabriella C. Russo, B. R. Si, Jungwoo Kim, Lauren Hoffner, Cholsoon Jang, Moonjung Jung, Robert D Leone, K. Konstantopoulos, Andrew J Ewald, Denis Wirtz, Sangmoo Jeong.(2024). E-cadherin Induces Serine Synthesis to Support Progression and Metastasis of Breast Cancer..
[29] I. O. Osman, Clémence Garrec, Gabriel Augusto Pires de Souza, A. Zarubica, Djamal Brahim Belhaouari, J. Baudoin, H. Lepidi, J. Mege, B. Malissen, B. Scola, C. Devaux.(2022). Control of CDH1/E-Cadherin Gene Expression and Release of a Soluble Form of E-Cadherin in SARS-CoV-2 Infected Caco-2 Intestinal Cells: Physiopathological Consequences for the Intestinal Forms of COVID-19.
[30] L. Thorstensen, G. E. Lind, T. Løvig, C. B. Diep, G. Meling, T. Rognum, R. Lothe.(2005). Genetic and epigenetic changes of components affecting the WNT pathway in colorectal carcinomas stratified by microsatellite instability..
[31] Hong Liu, Yan Ma, Hong-wei He, Wu-li Zhao, R. Shao.(2017). SPHK1 (sphingosine kinase 1) induces epithelial-mesenchymal transition by promoting the autophagy-linked lysosomal degradation of CDH1/E-cadherin in hepatoma cells.
[32] Xiao Tan, Wenbin Chen, D. Lv, Tao-wei Yang, Kai-hui Wu, Li-bin Zou, Junqi Luo, Xu-min Zhou, Guo-chang Liu, F. Shu, X. Mao.(2020). LncRNA SNHG1 and RNA binding protein hnRNPL form a complex and coregulate CDH1 to boost the growth and metastasis of prostate cancer.
[33] Yan He, Qimin Zhang, Yuan-Xiao Chen, Yingjian Wu, Yuan Quan, Weihua Chen, Jing Yao, Peijing Zhang.(2023). ZHX2 deficiency enriches hybrid MET cells through regulating E-cadherin expression.


Comments
Leave a Comment