Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-E04756m |
| Size | 96T,5×96T,10×96T |
| Price | Request a Quote |
| Trial Size |
24T ELISA Kit Trial Size (Only USD$150/ kit) * Sample kit cost can be deducted as a $30 credit for each 96-assay kit of the same analyte and brand you subsequently purchase within six months until depleted. More details >> Interested in a trial size? Please leave a message below.
|
| Have Questions? | Leave a Message or Start an on-line Chat |
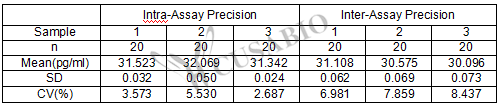
Intra-assay Precision (Precision within an assay): CV%<8%
Three samples of known concentration were tested twenty times on one plate to assess.
Inter-assay Precision (Precision between assays):CV%<10%
Three samples of known concentration were tested in twenty assays to assess.
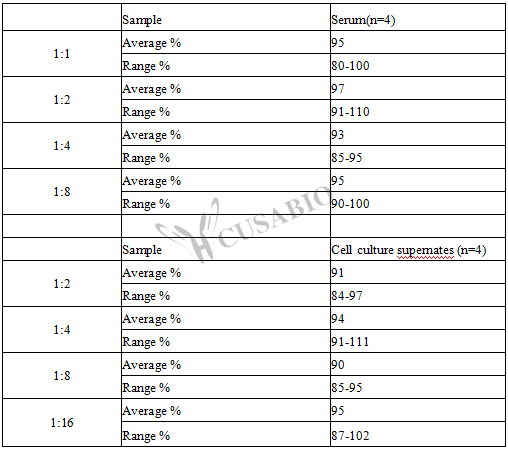
To assess the linearity of the assay, samples were spiked with high concentrations of Mouse VEGF in various matrices and diluted with the Sample Diluent to produce samples with values within the dynamic range of the assay.
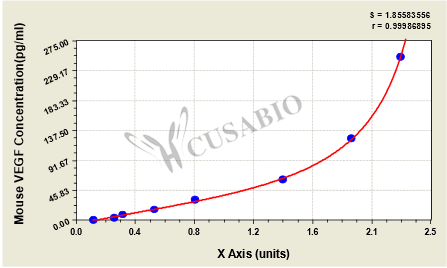
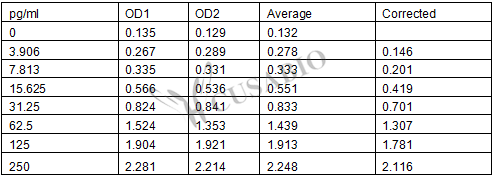
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed.
The mouse VEGFA ELISA Kit quantitates mouse VEGFA levels in multiple samples, including serum, plasma, cell culture supernates, and tissue homogenates. VEGFA usually referred to simply as VEGF, is a renowned angiogenic factor with potent mitogenic, pro-survival, anti-apoptotic, and vascular permeative properties. It plays an essential role in capillary maintenance, endothelial cell and myofiber survival, exercise-induced angiogenesis, and muscular endurance. VEGF binds to its receptor VEGFR1 and VEGFR2, activating several downstream signaling pathways, including MAPK and PI3K, that stimulate the expression of genes involving the promotion of endothelial cells' proliferation, migration, survival, and vascular permeability. VEGF-mediated signaling is associated with key aspects of tumorigenesis, including the function of cancer stem cells and tumor initiation.
This kit employs the sandwich-ELISA mechanism in conjugation with VEGFA antibody-VEGFA antigen-specific binding as well as HRP-TMB chromogenic reaction to measure the concentration of VEGFA in the samples. The kit is characterized by high sensitivity, strong specificity, good linearity, high recovery, and a precision of less than 10%.
Sample type: Other: Tumor interstitial fluid (TIF)
Sample species: Mouse
Sample dilution: 1:50
Review: I used CSB-E04756m to detect normal tumor interstitial fluid. The OD values of the standard curve are 0.106-1.566, and the OD values of the samples are 0.656-1.417.
By Anonymous
How to perform 2 freeze/thaw cycles in terms of freezing and thawing temperature and duration?
2 freeze/thaw cycles as follows: store the sample which was dealt with PBS homogenization overnight at -20°C, then allow the sample to dissolve thoroughly at room temperature. This is the first freeze/thaw. After that, freeze the sample at -80°C or store it overnight at -20°C, then allow the sample to dissolve thoroughly at room temperature again. This is the second freeze/thaw. by the way, the whole thaw procedure suggested being proceeded on the ice. For the succeeding procedures, you only need to follow the instruction.
Should we add protease and phosphatase inhibitors during Tissue homogenization?
If you want to deal with tissue homogenization, you should consider first the difference and homogeneity of the tissue sample. We suggest a preliminary experiment first with the gradient dilutions. Secondly, PBS will be used in the processing of tissue samples.
For CSB-E04756m, be careful not to add protease during tissue processing. This substance accelerates protein degradation. Phosphatase inhibitors can be added. The general ratio is 1:100. The specific dosage should be combined with the recommended dosage in the instructions of the phosphatase inhibitor.
What is the required PH of PBS used for tissue homogenization?
Usually,PBS will be used in the processing of tissue samples, pH should be 7.2-7.4.
Regarding calculations, should we normalize the results to the total protein content of each sample?
You could choose not to do the quantitation of the total protein. Just use pg/mL directly to express the result. You could also choose to do the quantitation of the total protein and then use pg/mg to express the result. It all depends on your request for the assay result.

