Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Nowadays, ample evidence has shown the roles of immune checkpoints in cancer immunotherapy. Intriguingly, CD226, a well-known active receptor, it is increasingly being recognized and researched through the two articles published on Oct. 13 in Immunity. One suggests that CD226 is absence in CD8+T cells as tumor infiltrates, and the loss of CD226 limits the checkpoint blockaded efficacy and CD137 agonists [1]. The other introduces that the degradation of CD226 induced by CD155 can enhance immune therapy resistance, whereas CD226 stimulation may represent a useful strategy to increase the efficacy immunotherapy [2].
The recent two researches have demonstrated that CD226 expression represents the important role for immune responses in cancer. CD226, as a focal point in cancer immunotherapy is making waves. In case you missed it, here are some highlights from CD226 studies to date.
1. CD226 Structure and Expression
3. CD226 Function on NK, CD8 + T cells, and APCs
4. CD226/TIGIT Pathway on T cells and APCs
CD226, namely Ig superfamily member DNAX accessory molecule 1 (DNAM-1), is an immunoglobulin superfamily member [3]. Human CD226 gene encodes a 67 kDa glycoprotein (336 amino acids) and is located on chromosome 18q22–23 with length of 2,487 bp [4]. An overview of CD226 molecular suggested that CD226 consists of three domains (Fig.1): the first domain of envelope is required for ligands recognition, adhesion, immune synapse formation and cytotoxic effect; the extracellular domain includes two Ig V-like domains and 8 N-linked glycoprotein sites; the intracellular domain contains four tyrosine residues and one serine residue. CD226 moves on the cell membrane as CD226 binds to its ligand, and then CD226 recruits intracellular signaling molecules such as PTK and PKC, which could activate the cells via phosphorylation on the four residues [5-6].
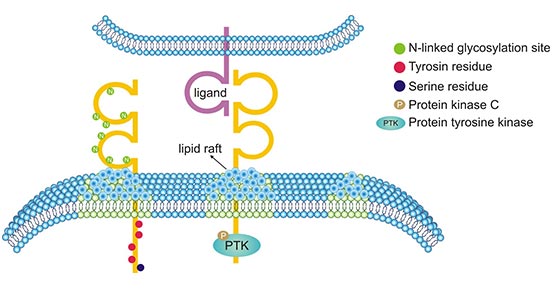
Figure 1. Three domains of CD226 molecular [6]
*This figure is derived from the publication on Frontiers in Cell and Developmental Biology [6]
In recent years, CD226 has been found widely expressed on T cells, NK cells, B cells, monocyte/macrophage, dendritic cells (DC), platelet lineage, and others [7]. It was also demonstrated that CD226 binding to its ligands cells could drive a series of regulation mechanisms in certain human tumors [8].
The poliovirus receptor (PVR; CD155) and its family member, poliovirus receptor-related 2 (Nectin-2; CD112), were identified as the ligands for CD226 by Bottino et al., in 2003 [9]. Both CD155 and CD112 belong to the members of the nicotine-like protein family and the nicotine protein family [10]. CD155 and CD112 molecules are widely expressed in a variety of tissue cells, such as nerve cells, endothelial cells, epithelial cells, antigen-presenting cells (APCs)/DC cells, fibroblasts, pathogen infected cells [11].
As the ligands of CD226, CD155 and CD112 expression affect biological functions of CD226, like Th1 differentiation and NK activation [12]. Besides, overexpressed CD155 and CD112 are found on multiple cancer cell lines and primary tumor [13]. Interactions between CD226 and the CD155/CD112 ligands play an important role in T cell and NK cell-mediated recognition and lysis of tumor cells.
CD226 is initially expressed on NK and T cells in human body as a functional protein [7]. Accumulating evidence shows that this adhesion molecule is closely related to the occurrence of a host of biological activities [14].
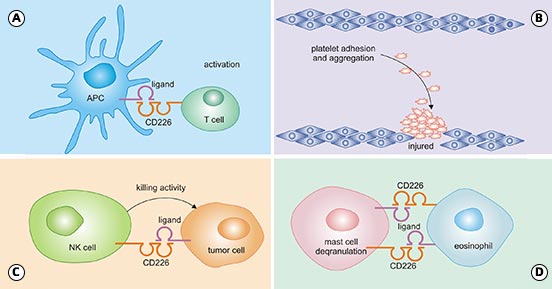
Figure 2. CD226 function on NK, CD8 + T cells, and APCs [6]
*This figure is derived from the publication on Frontiers in Cell and Developmental Biology [6]
The interaction of CD226/CD155 is very important for the interaction of CD8 + T cells and the immune response of APCs, which results in cell activation, proliferation, and differentiation (Fig.2A). In addition, CD226 influences the CD8+T cells migration to inflammatory site or tumor microenvironment: CD226 induces platelet activation and mediates platelet adhesion by cross-linking with its monoclonal antibodies (mAbs) (Fig.2B); furthermore, the CD226 activation via binding to CD155 and CD112, which triggers NK cells to kill tumor cells (Fig.2C); lastly, CD226 enhance the degranulation mediated by mast cell FCεRI, which is also probably achieved by the interaction of CD226/CD112 (Fig.2D) [6].
Recently a novel mechanism CD226/TIGIT that links TIGIT expression in the control of infections and malignancies has been identified [15]. TIGIT, a co-inhibitory receptor of the Ig-family acts as an important immune checkpoint in tumors [16]. Researchers found that the CD226/TIGIT regulates the function of T cells and APCs since they shared ligands CD155 and CD112 [17].
Using schematic illustration, the CD226/TIGIT pathway in the interaction between a T cell and APC was shown in brief. The T cell recognizes its cognate antigen in the context of MHC with its T cell receptor (TCR). For further activation, it needs co-stimulatory signals, which can be delivered via CD226 binding to its two ligands CD155 and CD112 expressed on the APC. CD155 and CD112 also could bind TIGIT. Meanwhile, CD155 is also the ligand of CD96 [18].
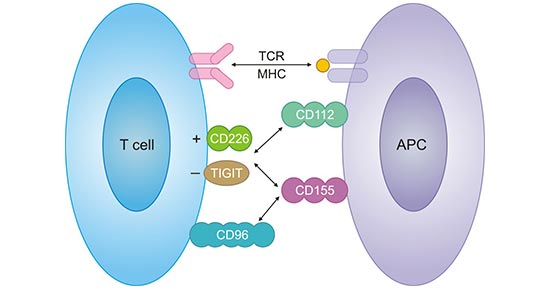
Figure 3. Schematic illustration of CD226/TIGIT pathway [18]
*This figure is derived from the publication on PLOS ONE [18]
Latest reports suggested that a high CD226/TIGIT expression in Tregs regulates their suppressive function and stability in tumors, and TIGIT exerts immune suppression effects by competing with CD226 for the same CD155 and CD112 ligand [15]. CD226/TIGIT suggested novel immunotherapies: to activate CD226 in T cells together with TIGIT blockade to counteract immune suppression, and finally to decrease tumor growth.
CD226 can work as an immune stimulator in anti-tumor immunity. a large number of studies have shown the prospect of CD226 in the anti-tumor immunity. On the one hand, CD226 has been found to regulate the NK cell cytotoxicity, stimulating NK cell-mediated killing of tumor cells, which indicates that CD226+NK cells play a crucial role in the recognition of human tumors [6, 19]. Another point is that many immune checkpoints have been identified in tumor, which either dampen the immune response (PD-L1, CTLA4) [18] or enhance the immune progress (CD28; CD137) [21-22].
Similarly, D226 can also work as a co-stimulatory receptor in anti-tumor immunity. its activation promotes anti-tumor immune response. For example, early research indicates that CD226 on hepatoma cells was down-regulated and its overexpression is related to the improved survival rate and survival time of patients [23]. A latest article suggests that CD226 agonist antibody-mediated activation of CD22 augments the effect of TIGIT blockade on CD8+ T cell responses, which further improve the immune efficacy in patients with pancreatic ductal adenocarcinoma [24].
Yet despite the good news, one question stands out, about clinical trials. In fact, today's global market for mAbs is highly unbalanced. A new analysis by the Associated Press has predicted that the antibody drug market will be worth more than USD240.2 billion by 2025 due to their growing popularity and therapeutic potential. With the advent of the bulk of immune checkpoints, numerous studies have focused on certain potential signal pathways to promote the immune response in tumor therapies.
Since further researches must wait until the trial has gathered more data, there is a large unmet need for CD226 antibody-based agents. Upon mAbs-mediated CD226, it drives activation of signals, which may attribute to tyrosine phosphorylation of CD226 itself, enhancement of cytotoxicity and cytokine production in T and NK cells. CD226, therefore, as a predictive biomarker for cancer immunotherapy, the engagement of CD226 could be a promising therapeutic strategy in cancer.
References
[1] Braun M, Aguilera AR, et al., CD155 on Tumor Cells Drives Resistance to Immunotherapy by Inducing the Degradation of the Activating Receptor CD226 in CD8+T Cells[J]. Immunity, 2020, 53(4): 805-823.e15.
[2] Weulersse M, Asrir A, et al., Eomes-Dependent Loss of the Co-activating Receptor CD226 Restrains CD8+ T Cell Anti-tumor Functions and Limits the Efficacy of Cancer Immunotherapy [J]. Immunity, 2020, 53(4): 824-839.e10.
[3] Nicolas Reymond, Anne-Marie Imbert, et al., DNAM-1 and PVR Regulate Monocyte Migration through Endothelial Junctions [J]. Journal of Experimental Medicine, 2004; 199(10): 1331–1341.
[4] Mattana T C C, Santos A S, Fukui R T, et al., CD226 rs763361 Is Associated with the Susceptibility to Type 1 Diabetes and Greater Frequency of GAD65 Autoantibody in a Brazilian Cohort[J]. Mediators of Inflammation, 2014.
[5] Han, Wang, Jianxun, et al., Binding mode of the side-by-side two-IgV molecule CD226/DNAM-1 to its ligand CD155/Necl-5[J]. Proceedings of the National Academy of Sciences of the United States of America, 2018.
[6] Huang Z, Qi G, Miller J S, et al., CD226: An Emerging Role in Immunologic Diseases [J]. Frontiers in Cell and Developmental Biology, 2020, 8.
[7] Kojima H, Kanada H, Shimizu S, et al., CD226 Mediates Platelet and Megakaryocytic Cell Adhesion to Vascular Endothelial Cells [J]. Journal of Biological Chemistry, 2003, 278(38): 36748-36753.
[8] Kim J S, Shin B R, Lee H K, et al., Cd226−/− natural killer cells fail to establish stable contacts with cancer cells and show impaired control of tumor metastasis in vivo[J]. OncoImmunology, 2017.
[9] Bottino C, Castriconi R, Pende D, et al., Identification of PVR (CD155) and Nectin-2 (CD112) as cell surface ligands for the human DNAM-1 (CD226) activating molecule [J]. Journal of Experimental Medicin, 2003, 198(4): 557-567.
[10] Tahara-Hanaoka S, Miyamoto A, Hara A, et al., Identification and characterization of murine DNAM-1 (CD226) and its poliovirus receptor family ligands [J]. Biochemical and Biophysical Research Communications, 2005, 329(3): 996-1000.
[11] Pende D, Castriconi R, Romagnani P, et al., Expression of the DNAM-1 ligands, Nectin-2 (CD112) and poliovirus receptor (CD155), on dendritic cells: relevance for natural killer-dendritic cell interaction [J]. Blood, 2006, 107(5): 2030-2036.
[12] Lozano E, Joller N, Cao Y, et al., The CD226/CD155 interaction regulates the proinflammatory (Th1/Th17)/anti-inflammatory (Th2) balance in humans [J]. Journal of Immunology, 2013, 191(7): 3673-3680.
[13] Iguchi-Manaka A, Okumura G, Ichioka E, et al., High expression of soluble CD155 in estrogen receptor-negative breast cancer [J]. Breast Cancer, 2020, 27(1): 92-99.
[14] Wagner A K, Kadri N, Sn?Ll J, et al., Expression of CD226 is associated to but not required for NK cell education[J]. Nature Communications, 2017, 8: 15627.
[15] Fourcade J, Sun Z, Chauvin JM, et al., CD226 opposes TIGIT to disrupt Tregs in melanoma [J]. JCI Insight, 2018, 26, 3(14): e121157.
[16] Chauvin JM, Zarour HM. TIGIT in cancer immunotherapy [J]. Immunother Cancer, 2020, 8(2): e000957.
[17] Lozano E, Dominguez-Villar M, Kuchroo V, et al., The TIGIT/CD226 axis regulates human T cell function[J]. Journal of Immunology, 2012, 188(8): 3869-3875.
[18] Kraus A K, Chen J, Edenhofer I, et al., The Role of T Cell Costimulation via DNAM-1 in Kidney Transplantation [J]. PLOS ONE, 2016.
[19] Du X, De Almeida P, Manieri N, et al., CD226 regulates natural killer cell antitumor responses via phosphorylation-mediated inactivation of transcription factor FOXO1[J]. Proceedings of the National Academy of Sciences, 2018.
[20] Brunner Weinzierl M C, Rudd C E. CTLA-4 and PD-1 Control of T-Cell Motility and Migration: Implications for Tumor Immunotherapy [J]. Frontiers in Immunology, 2018, 9.
[21] Waite JC, Wang B, Haber L, et al., Tumor-targeted CD28 bispecific antibodies enhance the antitumor efficacy of PD-1 immunotherapy [J]. Science Translational Medicine, 2020, 24, 12(549): eaba2325.
[22] Glorieux C, Huang P. CD137 expression in cancer cells: regulation and significance [J]. Cancer Communications, 2019, 39(1): 70.
[23] Gong J, Fang L, Liu R, et al., UPR decreases CD226 ligand CD155 expression and sensitivity to NK cell‐mediated cytotoxicity in hepatoma cells [J]. European Journal of Immunology, 2014, 44(12): 3758-67.
[24] Jin HS, Ko M, Choi DS, et al., CD226hiCD8+T Cells Are a Prerequisite for Anti-TIGIT Immunotherapy [J]. Cancer Immunol Res, 2020, 8(7): 912-925.


Comments
Leave a Comment