Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-E04650m |
| Size | 96T,5×96T,10×96T |
| Price | Request a Quote |
| Trial Size |
24T ELISA Kit Trial Size (Only USD$150/ kit) * Sample kit cost can be deducted as a $30 credit for each 96-assay kit of the same analyte and brand you subsequently purchase within six months until depleted. More details >> Interested in a trial size? Please leave a message below.
|
| Have Questions? | Leave a Message or Start an on-line Chat |
Intra-assay Precision (Precision within an assay): CV%<8% | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Three samples of known concentration were tested twenty times on one plate to assess. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Inter-assay Precision (Precision between assays): CV%<10% | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Three samples of known concentration were tested in twenty assays to assess. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
To assess the linearity of the assay, samples were spiked with high concentrations of mouse LEP in various matrices and diluted with the Sample Diluent to produce samples with values within the dynamic range of the assay. | ||||||||||||
| Sample | Serum(n=4) | ||||||||||
1:1 | Average % | 92 | ||||||||||
Range % | 86-95 | |||||||||||
1:2 | Average % | 103 | ||||||||||
Range % | 97-105 | |||||||||||
1:4 | Average % | 93 | ||||||||||
Range % | 87-95 | |||||||||||
1:8 | Average % | 96 | ||||||||||
Range % | 92-100 | |||||||||||
The recovery of mouse LEP spiked to levels throughout the range of the assay in various matrices was evaluated. Samples were diluted prior to assay as directed in the Sample Preparation section. | ||||||||||||
Sample Type | Average % Recovery | Range | ||||||||||
Serum (n=5) | 93 | 89-97 | ||||||||||
EDTA plasma (n=4) | 95 | 90-98 | ||||||||||
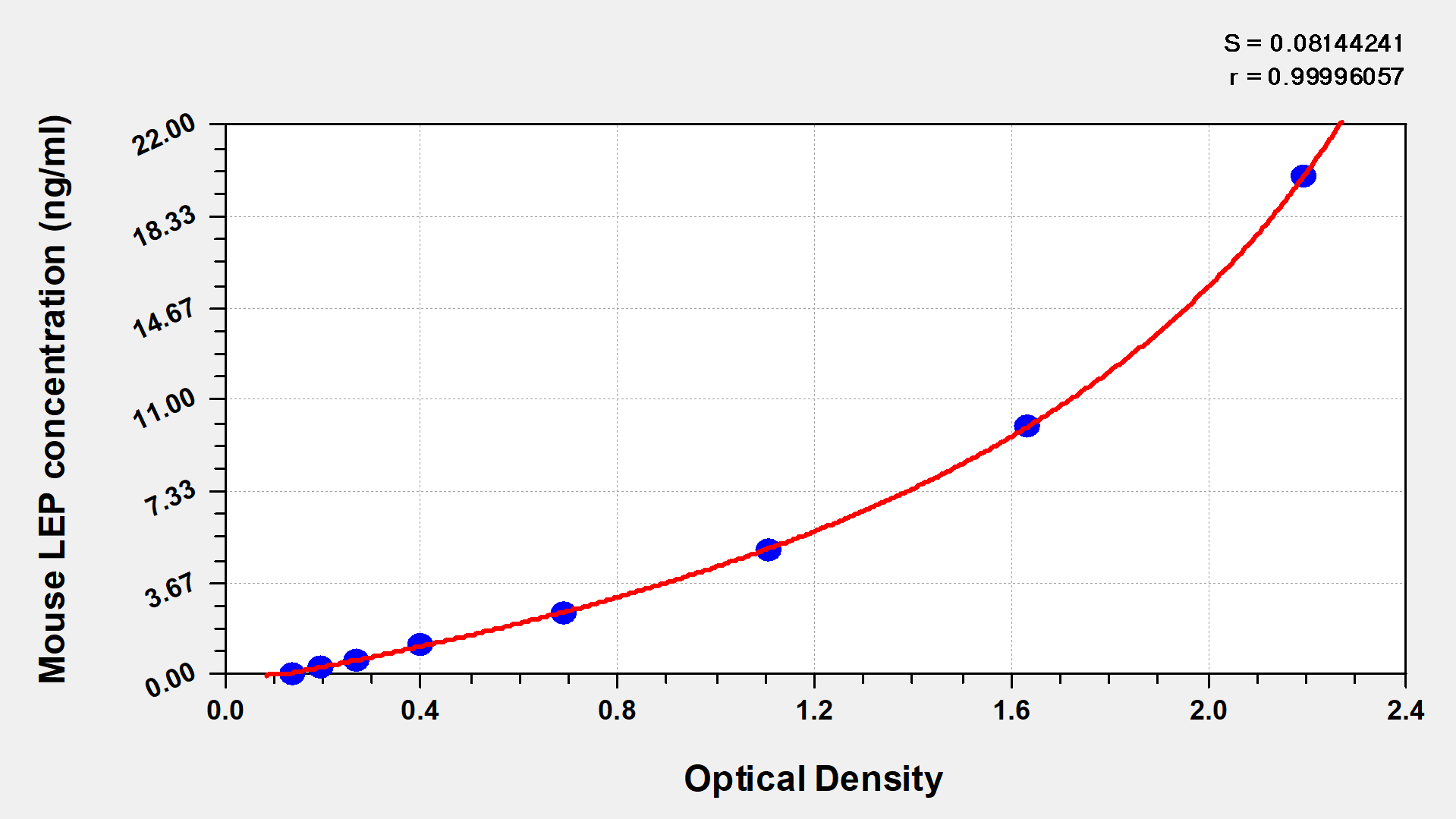
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed. | |||||||||||||
| |||||||||||||
ng/ml | OD1 | OD2 | Average | Corrected | |||||||||
20 | 2.196 | 2.184 | 2.190 | 2.040 | |||||||||
10 | 1.641 | 1.628 | 1.635 | 1.485 | |||||||||
5 | 1.118 | 1.103 | 1.111 | 0.961 | |||||||||
2.5 | 0.686 | 0.707 | 0.697 | 0.547 | |||||||||
1.25 | 0.415 | 0.403 | 0.409 | 0.259 | |||||||||
0.625 | 0.285 | 0.276 | 0.281 | 0.131 | |||||||||
0.312 | 0.202 | 0.214 | 0.208 | 0.058 | |||||||||
0 | 0.152 | 0.148 | 0.150 |
| |||||||||
This Mouse LEP ELISA Kit was designed for the quantitative measurement of Mouse LEP protein in serum, plasma, tissue homogenates. It is a Sandwich ELISA kit, its detection range is 0.312 ng/mL-20 ng/mL and the sensitivity is 0.078 ng/mL.
Sample type: Plasma (anticoagulant)
Sample species: Mouse
Sample dilution: No dilution
Review: I used E04650m to detect leptin index in mice. The standard curve OD value was 0.071, 0.119, 0.17, 0.218, 0.364, 0.574, 1.06, 1.523, and the sample OD value was about 0.062, which was lower than the detection limit
By Anonymous
Please could you tell us if you have a Mouse Leptin ELISA Kit that can
be used on only 5 ul of serum?

