Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-E07433r |
| Size | 96T,5×96T,10×96T |
| Price | Request a Quote |
| Trial Size |
24T ELISA Kit Trial Size (Only USD$150/ kit) * Sample kit cost can be deducted as a $30 credit for each 96-assay kit of the same analyte and brand you subsequently purchase within six months until depleted. More details >> Interested in a trial size? Please leave a message below.
|
| Have Questions? | Leave a Message or Start an on-line Chat |
Intra-assay Precision (Precision within an assay): CV%<8% | ||||||
Three samples of known concentration were tested twenty times on one plate to assess. | ||||||
Inter-assay Precision (Precision between assays): CV%<10% | ||||||
Three samples of known concentration were tested in twenty assays to assess. | ||||||
To assess the linearity of the assay, samples were spiked with high concentrations of rat LEP in various matrices and diluted with the Sample Diluent to produce samples with values within the dynamic range of the assay. | ||||||
| Sample | Serum(n=4) | ||||
1:1 | Average % | 88 | ||||
Range % | 80-92 | |||||
1:2 | Average % | 95 | ||||
Range % | 91-105 | |||||
1:4 | Average % | 94 | ||||
Range % | 89-97 | |||||
1:8 | Average % | 90 | ||||
Range % | 86-96 | |||||
The recovery of rat LEP spiked to levels throughout the range of the assay in various matrices was evaluated. Samples were diluted prior to assay as directed in the Sample Preparation section. | ||||||
Sample Type | Average % Recovery | Range | ||||
Serum (n=5) | 90 | 87-94 | ||||
EDTA plasma (n=4) | 99 | 94-107 | ||||
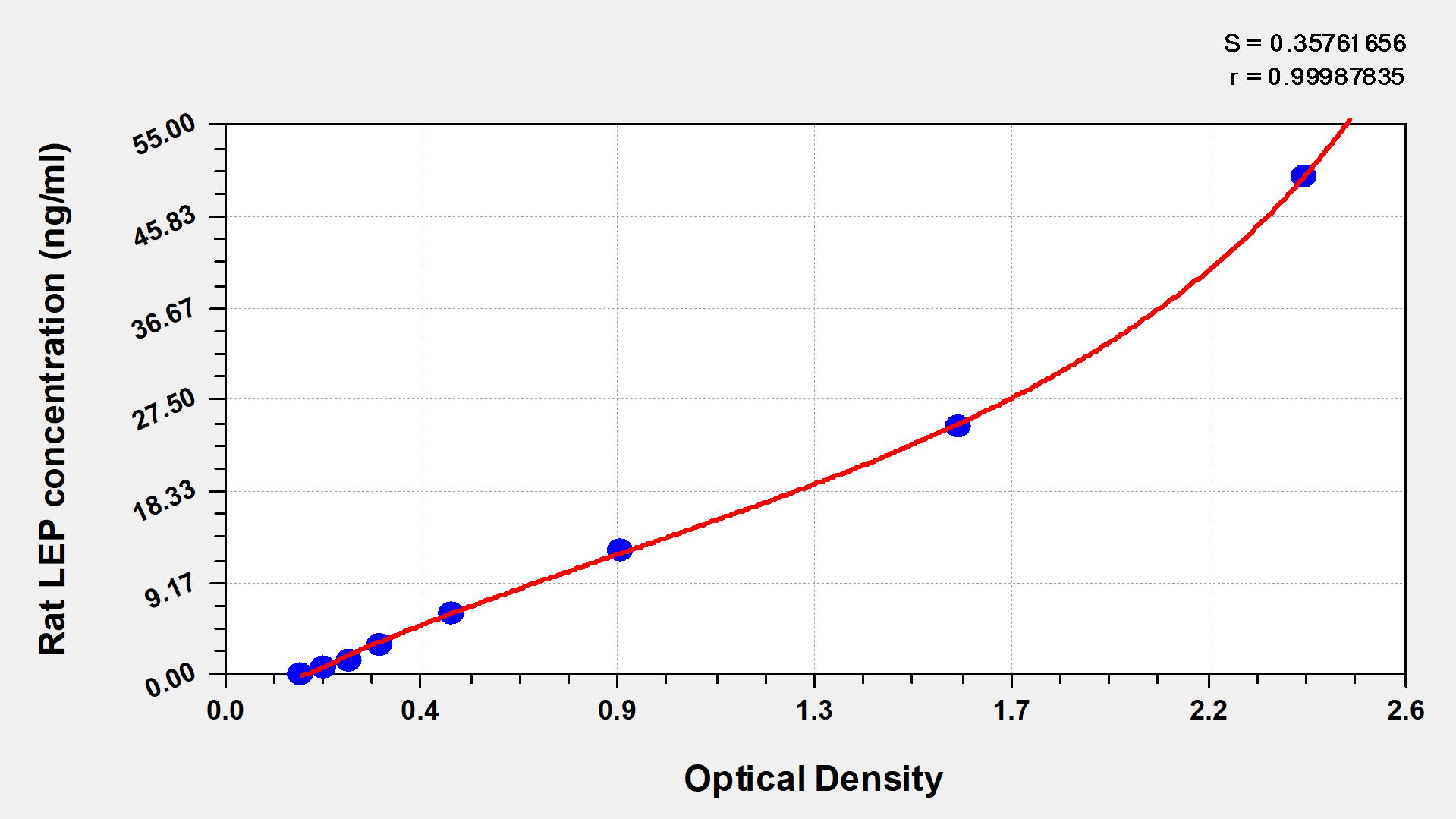
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed. | |||||||
| |||||||
ng/ml | OD1 | OD2 | Average | Corrected | |||
50 | 2.325 | 2.434 | 2.379 | 2.199 | |||
25 | 1.578 | 1.667 | 1.623 | 1.442 | |||
12.5 | 0.836 | 0.928 | 0.882 | 0.701 | |||
6.25 | 0.499 | 0.529 | 0.514 | 0.333 | |||
3.12 | 0.366 | 0.351 | 0.358 | 0.178 | |||
1.56 | 0.286 | 0.293 | 0.290 | 0.109 | |||
0.78 | 0.227 | 0.234 | 0.231 | 0.050 | |||
0 | 0.177 | 0.185 | 0.181 |
| |||
This rat Leptin (LEP) ELISA kit uses the quantitative sandwich enzyme immunoassay technique to measure the levels of mouse LEP in the samples, including serum, plasma, and tissue homogenates. Antibody specific for LEP has been pre-coated onto the microplate. Standards and samples are pipetted into the wells and any LEP present is bound by the immobilized antibody. After removing any unbound substances, a biotin-conjugated LEP antibody is added to the wells. After washing, avidin conjugated Horseradish Peroxidase (HRP) is added to the wells, forming an antibody-antigen-enzyme-labeled antibody complex. Following a wash to remove any unbound HRP-avidin, the TMB substrate solution is added to the wells and the color develops into blue. The color changes from blue to yellow after adding the stop solution to the wells. The color intensity is proportional to the amount of LEP bound in the initial step.
LEP is a hormone mainly synthesized by adipocytes and mainly acts on the brainstem and hypothalamus to regulate the long-term balance between the body's food intake and energy consumption as well as neuroendocrine function. LEP binding to LEPR activates and regulates several downstream signaling pathways, including JAK2/STAT3/5, MAPK/ERK, and PI3K pathways, contributing to LEP's anorexigenic effects. In addition to regulating energy homeostasis, LEP also plays several roles, including regulation of immune responses, supporting cell growth and tissue repair, and modulation of glucose and lipid metabolism. The deficiency of LEP is linked to dysregulation of cytokine production, enhanced susceptibility to infectious diseases, autoimmune disorders, malnutrition, and inflammatory responses.
There are currently no reviews for this product.

