Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
HRAS, a member of the RAS family alongside NRAS and KRAS, is encoded by the HRAS gene and functions as a membrane-associated GTP enzyme. This protein serves as a molecular switch, regulating cell growth, differentiation, and survival by binding to GTP or GDP, thus controlling cell signaling pathways. Abnormalities in HRAS, such as mutations or overexpression, are linked to the development of various cancers, including thyroid, salivary gland, bladder, cervical, and prostate cancers, highlighting its significant role in tumorigenesis.
HRAS Antibody (CSB-PA010726LA01HU)
Validated Data
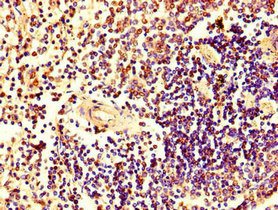
Immunohistochemistry of paraffin-embedded human spleen tissue using CSB-PA010726LA01HU at dilution of 1:100
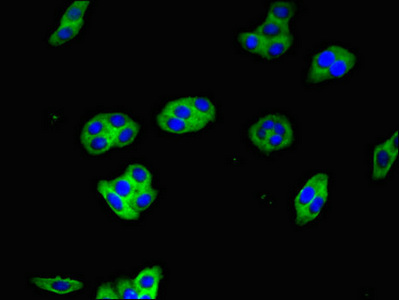
Immunofluorescent analysis of HepG2 cells using CSB-PA010726LA01HU at dilution of 1:100 and Alexa Fluor 488-congugated AffiniPure Goat Anti-Rabbit IgG(H+L)
The following HRAS reagents supplied by CUSABIO are manufactured under a strict quality control system. Multiple applications have been validated and solid technical support is offered.
HRAS Antibodies for Homo sapiens (Human)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA010726GA01HU | HRAS Antibody | Human,Mouse,Rat | ELISA,WB,IHC |
| CSB-PA064824 | HRAS Antibody | Human,Mouse,Rat | ELISA,WB |
| CSB-PA010726LA01HU | HRAS Antibody | Human | ELISA, IHC, IF |
| CSB-PA010726LB01HU | HRAS Antibody, HRP conjugated | Human | ELISA |
| CSB-PA010726LC01HU | HRAS Antibody, FITC conjugated | Human | |
| CSB-PA010726LD01HU | HRAS Antibody, Biotin conjugated | Human | ELISA |
HRAS Antibodies for Rattus norvegicus (Rat)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA15539D0Rb | Hras Antibody, Biotin conjugated | Rat | ELISA |
| CSB-PA15539A0Rb | Hras Antibody | Rat, Mouse, Human | ELISA, WB, IHC |
HRAS Proteins for Homo sapiens (Human)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP010726HU CSB-EP010726HU CSB-BP010726HU CSB-MP010726HU |
Recombinant Human GTPase HRas (HRAS) | Yeast E.coli Baculovirus Mammalian cell |
HRAS Proteins for Rattus norvegicus (Rat)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP321775RA CSB-EP321775RA CSB-BP321775RA CSB-MP321775RA CSB-EP321775RA-B |
Recombinant Rat GTPase HRas (Hras1) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
| CSB-RP155374r | Recombinant Rat GTPase HRas (Hras) | E.coli |
HRAS Proteins for Gallus gallus (Chicken)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP357443CH CSB-EP357443CH CSB-BP357443CH CSB-MP357443CH CSB-EP357443CH-B |
Recombinant Chicken GTPase HRas (HRAS1) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
HRAS Proteins for Mesocricetus auratus (Golden hamster)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP717104MRG CSB-EP717104MRG CSB-BP717104MRG CSB-MP717104MRG CSB-EP717104MRG-B |
Recombinant Mesocricetus auratus GTPase HRas (HRAS1) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
HRAS Proteins for Mus musculus (Mouse)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP730737MO CSB-EP730737MO CSB-BP730737MO CSB-MP730737MO CSB-EP730737MO-B |
Recombinant Mouse GTPase HRas (Hras1) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |

