Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Cathepsin K (encoded by CTSK) is a lysosomal cysteine protease, also known as cathepsin O2. It plays a core role in bone resorption: secreted by activated osteoclasts, it degrades type I collagen and elastin in the bone matrix, supporting bone remodeling. It acts as a downstream effector in the RANKL-RANK-OPG pathway, which governs osteoclast differentiation and activity.
Dysregulated CTSK is linked to osteoporosis (excessive bone loss), Paget’s disease of bone, and bone metastases from breast or prostate cancers. Drug development includes odanacatib, a phase III CTSK inhibitor for osteoporosis (withdrawn due to cardiovascular risks), plus preclinical candidates targeting bone-related disorders.
Recombinant Human Cathepsin K (CTSK) (CSB-YP006192HU)
Validated Data
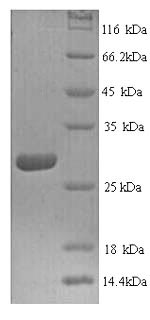
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
CTSK Antibody (CSB-PA006192ESR1HU)
Validated Data

Western blot
All lanes: CTSK antibody at 8μg/ml
Lane 1: MCF-7 whole cell lysate
Lane 2: Rat brain tissue
Secondary
Goat polyclonal to rabbit IgG at 1/10000 dilution
Predicted band size: 37 kDa
Observed band size: 37 kDa
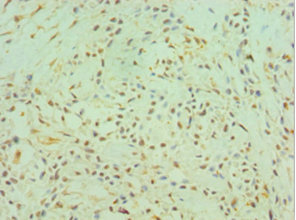
Immunohistochemistry of paraffin-embedded human breast cancer using CSB-PA006192ESR1HU at dilution of 1:100
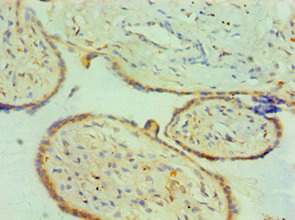
Immunohistochemistry of paraffin-embedded human placenta tissue using CSB-PA006192ESR1HU at dilution of 1:100
Human Cathepsin K,cath-K ELISA Kit (CSB-E09438h)
Validated Data
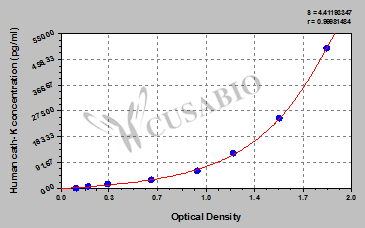
Code: CSB-E09438h
Size: 96T,5×96T,10×96T
Sensitivity: 1.95 pg/mL
Detection Range: 7.8 pg/mL-500 pg/mL
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed.
The following CTSK reagents supplied by CUSABIO are manufactured under a strict quality control system. Multiple applications have been validated and solid technical support is offered.
CTSK Antibodies for Rattus norvegicus (Rat)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA006192LA01RA | Ctsk Antibody | Rat | ELISA, IHC |
CTSK Antibodies for Homo sapiens (Human)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA006192GA01HU | CTSK Antibody | Human,Mouse,Rat | ELISA,WB,IHC |
| CSB-PA15119A0Rb | CTSK Antibody | Human | ELISA |
| CSB-PA006192ESR1HU | CTSK Antibody | Human, Rat | ELISA, WB, IHC |
| CSB-PA039740 | CTSK Antibody | Human,Mouse,Rat | ELISA,IHC |
| CSB-PA145433 | CTSK Antibody | Human,Mouse,Rat | ELISA,IHC |
CTSK Antibodies for Mus musculus (Mouse)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA006192ZA01MO | Ctsk Antibody | Mus musculus | ELISA, WB (ensure identification of antigen) |
CTSK Antibodies for Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA006192ZA01MOV | CTSK Antibody | Macaca fascicularis | ELISA, WB (ensure identification of antigen) |
CTSK Antibodies for Canis lupus familiaris (Dog) (Canis familiaris)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA006192ZA01DO | CTSK Antibody | Canis lupus familiaris | ELISA, WB (ensure identification of antigen) |
CTSK Antibodies for Oryctolagus cuniculus (Rabbit)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA006192ZA01RB | CTSK Antibody | Oryctolagus cuniculus | ELISA, WB (ensure identification of antigen) |
CTSK Proteins for Homo sapiens (Human)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192HU | Recombinant Human Cathepsin K (CTSK) | Yeast |
| CSB-BP006192HU CSB-MP006192HU CSB-EP006192HU-B |
Recombinant Human Cathepsin K (CTSK) | Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
| CSB-EP006192HU | Recombinant Human Cathepsin K (CTSK) | E.coli |
CTSK Proteins for Oryctolagus cuniculus (Rabbit)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192RB CSB-BP006192RB CSB-MP006192RB CSB-EP006192RB-B |
Recombinant Rabbit Cathepsin K (CTSK) | Yeast Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
| CSB-EP006192RB | Recombinant Rabbit Cathepsin K (Ctsk) | E.coli |
CTSK Proteins for Mus musculus (Mouse)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192MO CSB-BP006192MO CSB-MP006192MO CSB-EP006192MO-B |
Recombinant Mouse Cathepsin K (Ctsk) | Yeast Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
| CSB-EP006192MO | Recombinant Mouse Cathepsin K (Ctsk) | E.coli |
CTSK Proteins for Macaca mulatta (Rhesus macaque)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192MOW CSB-EP006192MOW CSB-BP006192MOW CSB-MP006192MOW CSB-EP006192MOW-B |
Recombinant Macaca mulatta (Rhesus macaque) Cathepsin K (CTSK) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
CTSK Proteins for Rattus norvegicus (Rat)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192RA CSB-BP006192RA CSB-MP006192RA CSB-EP006192RA-B |
Recombinant Rat Cathepsin K (Ctsk) | Yeast Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
| CSB-EP006192RA | Recombinant Rat Cathepsin K (Ctsk) | E.coli |
CTSK Proteins for Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192MOV CSB-BP006192MOV CSB-MP006192MOV CSB-EP006192MOV-B |
Recombinant Macaca fascicularis Cathepsin K (CTSK) | Yeast Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
| CSB-EP006192MOV | Recombinant Macaca fascicularis Cathepsin K (CTSK) | E.coli |
CTSK Proteins for Gallus gallus (Chicken)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192CH CSB-EP006192CH CSB-BP006192CH CSB-MP006192CH CSB-EP006192CH-B |
Recombinant Chicken Cathepsin K (CTSK) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
CTSK Proteins for Sus scrofa (Pig)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192PI CSB-EP006192PI CSB-BP006192PI CSB-MP006192PI CSB-EP006192PI-B |
Recombinant Pig Cathepsin K (CTSK) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
CTSK Proteins for Canis lupus familiaris (Dog) (Canis familiaris)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192DO CSB-BP006192DO CSB-MP006192DO CSB-EP006192DO-B |
Recombinant Dog Cathepsin K (CTSK) | Yeast Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
| CSB-EP006192DO | Recombinant Dog Cathepsin K (CTSK) | E.coli |
CTSK Proteins for Bos taurus (Bovine)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006192BO CSB-EP006192BO CSB-BP006192BO CSB-MP006192BO CSB-EP006192BO-B |
Recombinant Bovine Cathepsin K (CTSK) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
CTSK ELISA Kit for Homo sapiens (Human)
| Code | Product Name | Sample Types | Sensitivity |
|---|---|---|---|
| CSB-E09438h | Human Cathepsin K,cath-K ELISA Kit | serum, plasma, tissue homogenates | 1.95 pg/mL |
CTSK ELISA Kit for Mus musculus (Mouse)
| Code | Product Name | Sample Types | Sensitivity |
|---|---|---|---|
| CSB-E09440m | Mouse Cathepsin K,cath-K ELISA Kit | serum, plasma, tissue homogenates | 3.9 pg/mL |
CTSK ELISA Kit for Rattus norvegicus (Rat)
| Code | Product Name | Sample Types | Sensitivity |
|---|---|---|---|
| CSB-E09439r | Rat Cathepsin K,cath-K ELISA Kit | serum, plasma, tissue homogenates | 3.9 pg/mL |

