Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
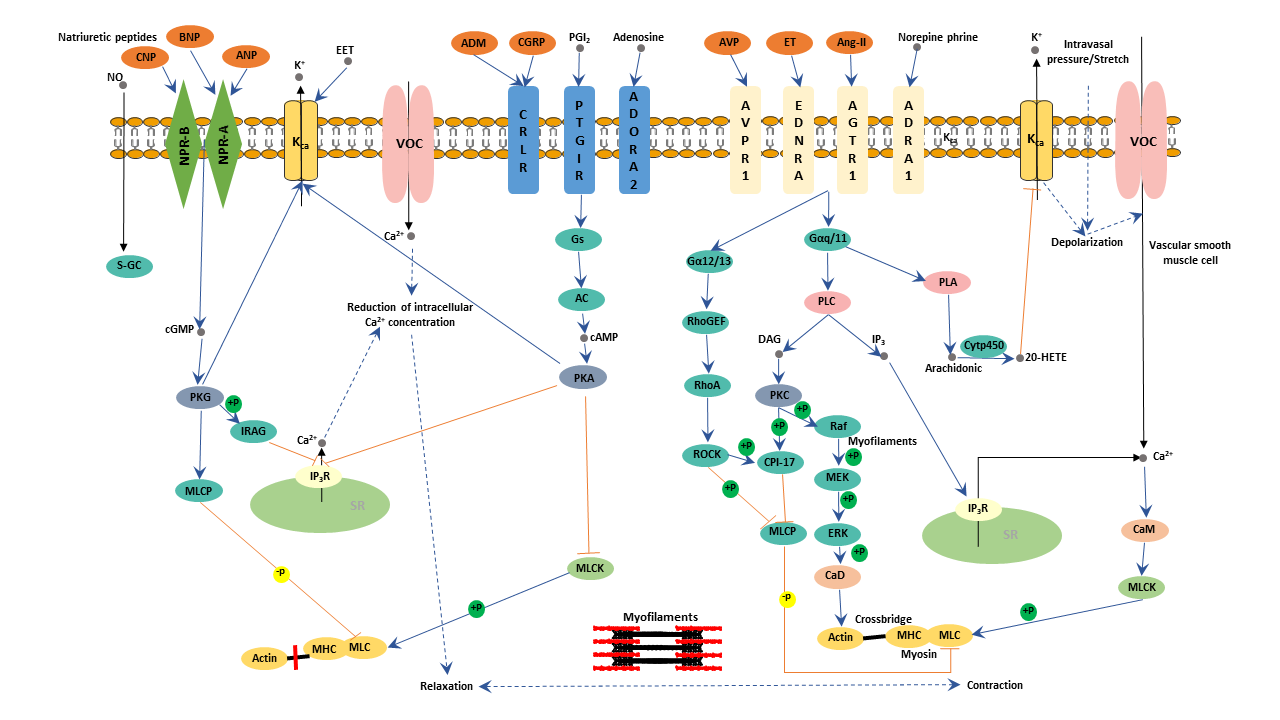
Contractile stimuli such as mechanical, electrical, and chemical stimuli leads to an increase in calcium concentration, which causes calcium influx to vascular smooth muscle cells and then triggers vasoconstriction.
Vascular smooth muscle contraction is a slow, sustained, and tonic process, whereas cardiac muscle undergoes a fast and relative short-cycle contraction so that the heart's indoor pressure rapidly increases in a short time.
Both cardiomyocytes and vascular smooth muscle contraction are dependent on the hydrolysis of ATP by the ATP enzyme in the myosin head to provide energy for the transverse bridge to slide and then tract thin muscle filament. But their trigger mechanisms are different. In cardiomyocytes, Ca2+ binds to troponin to stimulate the contraction process, whereas vascular smooth muscle contraction occurs through the binding and activation of Ca2+ to MLCK (myosin light chain kinase) via CaM (calmodulin).
The contraction or relaxation of vascular smooth muscle cells alter blood vessel volume and local blood pressure, and they both synergize to redistribute blood to where oxygen consumption is temporarily increased. Excessive vasoconstriction causes hypertension, while excessive vasodilation as in shock leads to hypotension.
There are two synergistic regulation pathways of vascular smooth muscle contraction, including calcium-dependent and calcium-independent regulation.
Calcium-dependent regulation is associated with transient phasic contraction. Stimulation by a large number of G-protein coupled receptor-mediated agonists such as alpha agonists or by depolarization of the cell membrane by channel activity or experimentally by equimolar replacement of NaCl with KCl in physiologic saline solution, cytoplasmic Ca2+ levels increase. All these eventually trigger extracellular Ca2+ influx into the cytoplasm through Ca2+ channels or the release of sarcoplasmic reticulum (SR) to facilitate intracellular Ca2+ increase.
As intracellular Ca2+ concentration rises, Ca2+ binds to CaM to form Ca2+/CaM complex. The Ca2+/CaM complex binds to and activates the MLCK. Activated MLCK phosphorylates myosin light chain (MLC) into MLC-p. Myosin subsequently combines with actin and evokes the activation of actin ATPase, triggering vascular smooth muscle contraction.
The calcium-independent pathway, also called Rho/Rho-associated protein kinase pathway, is related to the prolonged tonic contraction of vascular smooth muscle cells.
The calcium-independent pathway involves Rho/Rho-associated protein kinase (ROCK). Like all other Ras, RhoA is a GTPase. This means RhoA can act as a molecular switch between a GTP/GDP bound state. Normally, RhoA is bound to GDP to form an inactive RhoA/GDP. When stimulated by GPCR receptors, RhoA is activated by binding to RhoGTPase guanine nucleotide exchange factors. Active RhoA/GTP combines with and then activates its target proteins such as ROCK. Vascular smooth muscle cells express both ROCK1 and ROCK2. Activated ROCK exerts its role in vascular smooth muscle cells in two ways. One way is that ROCK regulates cytoskeletal organization by blocking actin filament depolymerization. Additionally, ROCK inhibits MLCP (myosin light-chain phosphatase). MLCP is an enzyme that dephosphorylates MLC. Therefore, MLCP inhibition keeps myosin light chain phosphorylated, which stimulates vascular smooth muscle contraction and increases vascular tone.
Recent research indicates that sphingosine-1-phosphate (S1P) signaling is an important regulator of vascular smooth muscle contraction. When transmural pressure increases, sphingosine kinase 1 (SPK1) phosphorylates sphingosine to S1P, which binds to the S1P2 receptor on the plasma membrane of cells. This results in a transient increase in intracellular Ca2+, activating Rac and Rhoa signaling pathways. Collectively, all these increase MLCK activity and decrease MLCP activity, promoting muscle contraction. This also allows arterioles to increase the resistance of increased blood pressure and is conducive to maintain constant blood flow.

