Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
BCL2L1, B-cell lymphoma/leukemia 2-like protein 1, alias BCL2L01, Bcl-2-related protein A1, Bfl-1, etc. BCL2L1 is an anti-apoptotic protein, a member of the Bcl-2 family, encoded by its own gene in human cells. It plays an important role in the regulation of apoptosis and inhibits the mitochondria-mediated apoptotic pathway mainly by interacting with pro-apoptotic proteins such as Bax or Bak.The mechanism of action of BCL2L1 involves its dynamic distribution between the cytoplasm and mitochondria, as well as its modulation of the permeability of the mitochondrial membrane.
The biological significance of BCL2L1 is very broad, and it participates in cellular response to a variety of stresses It is involved in cellular adaptation to a variety of stress responses, including oxidative stress, DNA damage, cytokine-induced stress, etc. BCL2L1 expression is regulated by a variety of signaling pathways, such as p53, NF-κB, MAPK, etc., which play a key role in the balance between cell survival and apoptosis.Abnormal BCL2L1 function is associated with the development of a variety of diseases, especially closely related to the development of tumors. In tumor cells, overexpression of BCL2L1 contributes to the escape of tumor cells from immune surveillance and chemotherapy-induced apoptosis, making it a potential target in tumor therapy.
Recombinant Human Bcl-2-like protein 1 (BCL2L1), partial (CSB-EP002613HU)
Validated Data
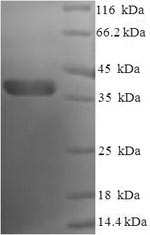
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
BCL2L1 Recombinant Monoclonal Antibody (CSB-RA247982A0HU)
Validated Data

Western Blot
Positive WB detected in: K562 whole cell lysate, Hela whole cell lysate, NIH/3T3 whole cell lysate, Rat Brain whole cell lysate, Mouse Brain whole cell lysate
All lanes: Bcl-XL antibody at 1:1000
Secondary
Goat polyclonal to rabbit IgG at 1/50000 dilution
Predicted band size: 27, 19, 26 kDa
Observed band size: 30 kDa
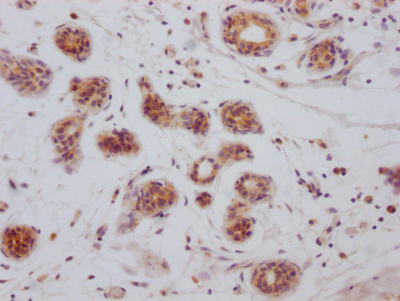
IHC image of CSB-RA247982A0HU diluted at 1:100 and staining in paraffin-embedded human breast cancer performed on a Leica BondTM system. After dewaxing and hydration, antigen retrieval was mediated by high pressure in a citrate buffer (pH 6.0). Section was blocked with 10% normal goat serum 30min at RT. Then primary antibody (1% BSA) was incubated at 4℃ overnight. The primary is detected by a Goat anti-rabbit IgG polymer labeled by HRP and visualized using 0.05% DAB.
Human Bcl-2-like protein 1(BCL2L1) ELISA kit (CSB-EL002613HU)
Validated Data
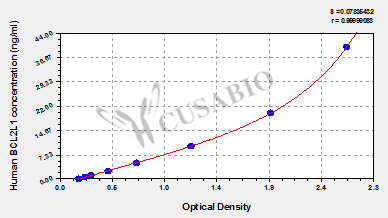
Code: CSB-EL002613HU
Size: 96T,5×96T,10×96T
Sensitivity: 0.156 ng/mL
Detection Range: 0.625 ng/mL-40 ng/mL
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed.
The following BCL2L1 reagents supplied by CUSABIO are manufactured under a strict quality control system. Multiple applications have been validated and solid technical support is offered.
BCL2L1 Antibodies for Homo sapiens (Human)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA027199GA01HU | BCL2L1 Antibody | Human,Mouse,Rat | ELISA,WB,IHC |
| CSB-MA0026131A0m | BCL2L1 Monoclonal Antibody | Human | ELISA,WB,IHC |
| CSB-PA002613HA01HU | BCL2L1 Antibody | Human, Mouse | ELISA, WB, IHC, IF |
| CSB-PA002613HB01HU | BCL2L1 Antibody, HRP conjugated | Human | ELISA |
| CSB-PA923588 | BCL2L1 (Ab-62) Antibody | Human,Mouse,Rat | ELISA,WB |
| CSB-PA786016 | Phospho-BCL2L1 (Ser62) Antibody | Human, Mouse, Rat | ELISA, WB, IHC, IF |
| CSB-PA000986 | BCL2L1 Antibody | Human,Mouse,Rat | WB, IHC, IF, ELISA |
| CSB-PA000987 | BCL2L1 Antibody | Human,Mouse,Rat | WB, IHC, ELISA |
| CSB-PA040205 | Phospho-BCL2L1 (S62) Antibody | Human,Mouse,Rat | WB, IHC, ELISA |
| CSB-PA040206 | Phospho-BCL2L1 (T47) Antibody | Human,Mouse,Rat | WB, IHC, IF, ELISA |
| CSB-PA040207 | Phospho-BCL2L1 (T115) Antibody | Human,Mouse,Rat | IHC, ELISA |
| CSB-PA040208 | BCL2L1 Antibody | Human,Mouse,Rat | IHC, ELISA |
| CSB-PA080279 | BCL2L1 Antibody | Human,Mouse,Rat | IHC |
| CSB-PA223208 | BCL2L1 Antibody | Human,Mouse,Rat | ELISA,WB |
| CSB-PA315068 | BCL2L1 Antibody | Human,Mouse,Rat | ELISA,WB |
| CSB-RA247982A0HU | BCL2L1 Recombinant Monoclonal Antibody | Human, Mouse, Rat | ELISA, WB, IHC |
BCL2L1 Proteins for Homo sapiens (Human)
| Code | Product Name | Source |
|---|---|---|
| CSB-EP002613HU | Recombinant Human Bcl-2-like protein 1 (BCL2L1), partial | E.coli |
| CSB-YP002613HU CSB-BP002613HU CSB-MP002613HU CSB-EP002613HU-B |
Recombinant human Bcl-2-like protein 1 | Yeast Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
BCL2L1 Proteins for Mus musculus (Mouse)
| Code | Product Name | Source |
|---|---|---|
| CSB-CF731050MO | Recombinant Mouse Bcl-2-like protein 1 (Bcl2l1) | in vitro E.coli expression system |
| CSB-YP731050MO1 CSB-EP731050MO1 CSB-BP731050MO1 CSB-MP731050MO1 CSB-EP731050MO1-B |
Recombinant Mouse Bcl-2-like protein 1 (Bcl2l1), partial | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
BCL2L1 Proteins for Rattus norvegicus (Rat)
| Code | Product Name | Source |
|---|---|---|
| CSB-CF002613RA | Recombinant Rat Bcl-2-like protein 1 (Bcl2l1) | in vitro E.coli expression system |
| CSB-YP002613RA1 CSB-EP002613RA1 CSB-BP002613RA1 CSB-MP002613RA1 CSB-EP002613RA1-B |
Recombinant Rat Bcl-2-like protein 1 (Bcl2l1), partial | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
BCL2L1 Proteins for Sus scrofa (Pig)
| Code | Product Name | Source |
|---|---|---|
| CSB-CF002613PI | Recombinant Pig Bcl-2-like protein 1 (BCL2L1) | in vitro E.coli expression system |
| CSB-YP002613PI1 CSB-EP002613PI1 CSB-BP002613PI1 CSB-MP002613PI1 CSB-EP002613PI1-B |
Recombinant Pig Bcl-2-like protein 1 (BCL2L1), partial | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
BCL2L1 ELISA Kit for Homo sapiens (Human)
| Code | Product Name | Sample Types | Sensitivity |
|---|---|---|---|
| CSB-EL002613HU | Human Bcl-2-like protein 1(BCL2L1) ELISA kit | serum, plasma, tissue homogenates | 0.156 ng/mL |
BCL2L1 ELISA Kit for Rattus norvegicus (Rat)
| Code | Product Name | Sample Types | Sensitivity |
|---|---|---|---|
| CSB-E13604r | Rat Bcl-2-like protein 1(Bcl2-L-1/Bcl-X)ELISA Kit | serum, plasma, tissue homogenates, cell lysates | 11.72 pg/mL |

