Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Cathepsin C (CTSC), also called dipeptidyl peptidase I (DPP-I), is a lysosomal cysteine protease expressed in innate immune cells like neutrophils and macrophages. It activates proforms of serine proteases (neutrophil elastase, cathepsin G) via dipeptidyl cleavage, critical for host defense and inflammatory responses. This links CTSC to pathways regulating neutrophil degranulation and pathogen clearance.
Biallelic CTSC mutations cause Papillon-Lefèvre syndrome (PLS, palmoplantar keratoderma + early periodontitis) and Haim-Munk syndrome (PLS variant with arachnodactyly). Selective inhibitors like brensocatib (phase3 trials) target CTSC to reduce neutrophil-driven inflammation, evaluated for non-cystic fibrosis bronchiectasis and COPD to mitigate exacerbations.
Recombinant Human Dipeptidyl peptidase 1 (CTSC) (Active) (CSB-MP006186HU)
Validated Data
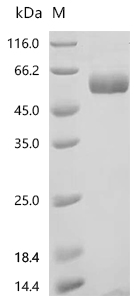
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
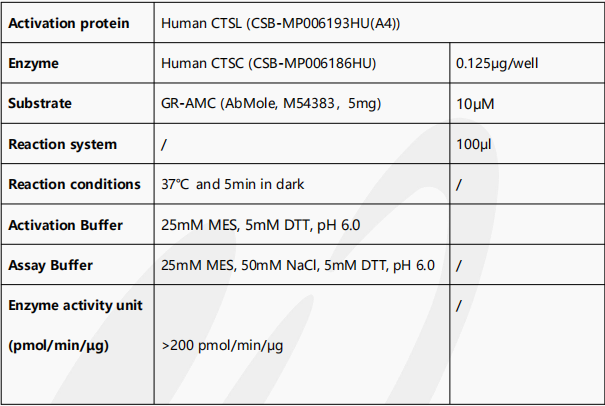
Activity
Measured by its ability to cleave the fluorogenic peptide substrate, Gly-Arg-7-amido-4-methylcoumarin (GR-AMC). The specific activity is >200 pmol/min/μg. The Human CTSC needs to be activated by Human CTSL (CSB-MP006193HU(A4)).
CTSC Antibody (CSB-PA006186LA01HU)
Validated Data
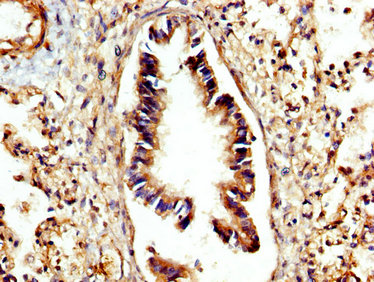
IHC image of CSB-PA006186LA01HU diluted at 1:300 and staining in paraffin-embedded human lung tissue performed on a Leica BondTM system. After dewaxing and hydration, antigen retrieval was mediated by high pressure in a citrate buffer (pH 6.0). Section was blocked with 10% normal goat serum 30min at RT. Then primary antibody (1% BSA) was incubated at 4°C overnight. The primary is detected by a biotinylated secondary antibody and visualized using an HRP conjugated SP system.
The following CTSC reagents supplied by CUSABIO are manufactured under a strict quality control system. Multiple applications have been validated and solid technical support is offered.
CTSC Antibodies for Homo sapiens (Human)
| Code | Product Name | Species Reactivity | Application |
|---|---|---|---|
| CSB-PA000029 | Cleaved-CTSC (R394) Antibody | Human | WB, ELISA |
| CSB-PA115010 | CTSC Antibody | Human | ELISA,WB,IHC |
| CSB-PA358929 | CTSC Antibody | Human | ELISA,WB,IHC |
| CSB-PA006186LA01HU | CTSC Antibody | Human | ELISA, IHC |
| CSB-PA006186LB01HU | CTSC Antibody, HRP conjugated | Human | ELISA |
| CSB-PA006186LC01HU | CTSC Antibody, FITC conjugated | Human | |
| CSB-PA006186LD01HU | CTSC Antibody, Biotin conjugated | Human | ELISA |
CTSC Proteins for Rattus norvegicus (Rat)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006186RA CSB-EP006186RA CSB-BP006186RA CSB-MP006186RA CSB-EP006186RA-B |
Recombinant Rat Dipeptidyl peptidase 1 (Ctsc) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
CTSC Proteins for Mus musculus (Mouse)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006186MO CSB-BP006186MO CSB-MP006186MO CSB-EP006186MO-B |
Recombinant Mouse Dipeptidyl peptidase 1 (Ctsc), partial | Yeast Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
CTSC Proteins for Homo sapiens (Human)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006186HU CSB-EP006186HU CSB-BP006186HU CSB-EP006186HU-B |
Recombinant Human Dipeptidyl peptidase 1 (CTSC), partial | Yeast E.coli Baculovirus In Vivo Biotinylation in E.coli |
| CSB-MP006186HU | Recombinant Human Dipeptidyl peptidase 1 (CTSC) (Active) | Mammalian cell |
CTSC Proteins for Canis lupus familiaris (Dog) (Canis familiaris)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP006186DO CSB-EP006186DO CSB-BP006186DO CSB-MP006186DO CSB-EP006186DO-B |
Recombinant Dog Dipeptidyl peptidase 1 (CTSC) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
CTSC Proteins for Bos taurus (Bovine)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP671368BO CSB-EP671368BO CSB-BP671368BO CSB-MP671368BO CSB-EP671368BO-B |
Recombinant Bovine Dipeptidyl peptidase 1 (CTSC) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
CTSC Proteins for Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP717180MOV CSB-EP717180MOV CSB-BP717180MOV CSB-MP717180MOV CSB-EP717180MOV-B |
Recombinant Macaca fascicularis Dipeptidyl peptidase 1 (CTSC) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |
CTSC Proteins for Pongo abelii (Sumatran orangutan) (Pongo pygmaeus abelii)
| Code | Product Name | Source |
|---|---|---|
| CSB-YP735773PYX CSB-EP735773PYX CSB-BP735773PYX CSB-MP735773PYX CSB-EP735773PYX-B |
Recombinant Pongo abelii Dipeptidyl peptidase 1 (CTSC) | Yeast E.coli Baculovirus Mammalian cell In Vivo Biotinylation in E.coli |

