Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-AP003861HU |
| Abbreviation | Recombinant Human TGFB1 protein, partial (Active) |
| MSDS | |
| Size | $290 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
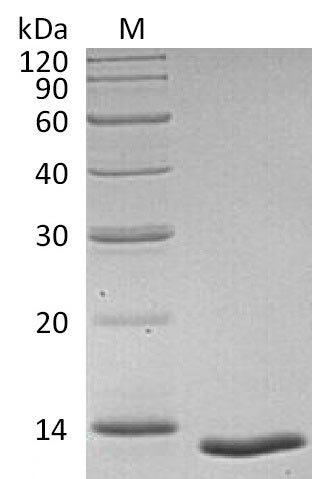
TGF-β1 serves as a master regulator of immune suppression and tumor microenvironment remodeling, making precise bioactivity essential for meaningful in vitro and in vivo cancer biology studies. This tag-free recombinant human TGF-β1, spanning the mature signaling domain (residues 279–390), demonstrates potent receptor-binding activity with an ED50 of 4–40 pg/mL in inhibiting IL-4-dependent TF-1 cell proliferation—a level of potency that supports use in cell proliferation assays, receptor-ligand binding studies via SPR or ELISA, and tumor biology models examining immune evasion or epithelial-mesenchymal transition. Mammalian cell expression preserves native disulfide bonding and folding critical for proper TGF-β receptor engagement, while endotoxin levels below 1.0 EU/μg combined with greater than 95% purity satisfy the criteria typically required for sensitive cell-based differentiation assays and antibody characterization workflows where background interference must remain minimal.
Applications : Stem/immune cell maintenance or differentiation
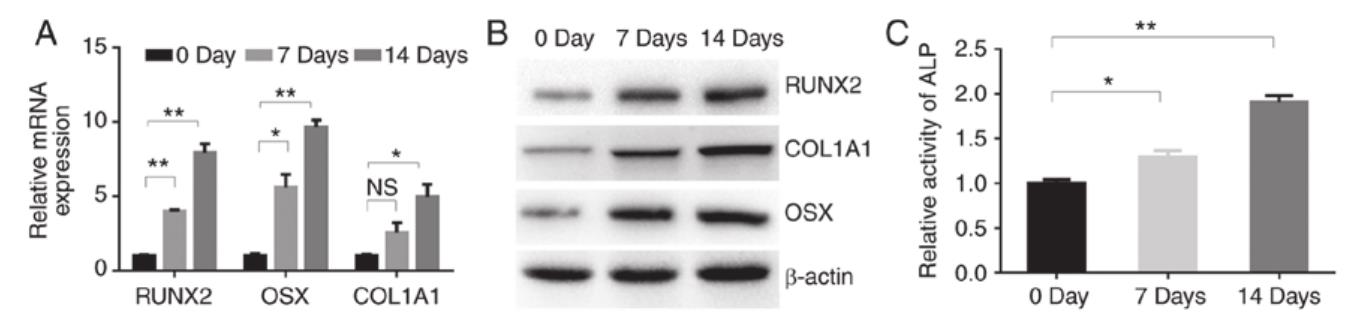
Review: TGF- β 1 promotes the osteoblastic differentiation of MC3T3-E1 cells. (A) RT-qPCR analysis of the osteogenic-related genes RUNX2, OSX and COL1A1 in MC3T3-E1 cells after treatment with TGF- β 1 (4 ng/ml) for 0, 7 and 14 days. (B) Western blotting of RUNX2, COL1A1‑and OSX protein in MC3T3-E1 cells cultured with TGF- β 1 (4 ng/ml) for 0, 7 and 14 days. (C) ALP activity in MC3T3‑E1 cells after treatment with TGF‑ β 1 (4 ng/ml) for 0, 7 and 14 days. NS, no significance change; * P<0.05, ** P<0.01 as indicated. TGF, transforming growth factor; RUNX2, runt-related transcription factor 2; OSX, osterix; COL1A1 collagen type I α 1 chain; ALP, alkaline phosphatase.
By Anonymous

