Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
CD70 (CD27L/TNFSF7), a member of the tumor necrosis factor receptor superfamily, has demonstrated significant potential in cancer immunotherapy and autoimmune disease research in recent years. Its widespread expression across various solid tumors and hematologic malignancies, coupled with restricted expression in normal tissues, positions it as an ideal therapeutic target. However, the heterogeneous expression of CD70 in solid tumors has long been a critical bottleneck limiting the efficacy of CAR-T and other cell-based therapies.
A groundbreaking study recently published in Science has challenged conventional understanding: so-called "antigen-negative" tumor cells have not actually lost CD70 expression entirely. Instead, its expression is suppressed to ultra-low levels through EZH2-mediated H3K27me3 epigenetic modification, creating a state of "pseudo-heterogeneity." Based on this discovery, researchers developed high-sensitivity HIT receptor CAR-T cells (CD70-HIT-T) capable of recognizing trace amounts of CD70, achieving complete tumor clearance in models of renal cancer, ovarian cancer, and pancreatic cancer.
Meanwhile, the domestic cell therapy landscape surrounding the CD70 target has become increasingly diversified, with multiple technological approaches including CAR-T, CAR-NK, and CAR-NKT advancing in parallel. Several products have already entered clinical stages and demonstrated preliminary efficacy. This article will explore the prospects and challenges of this target in solid tumor treatment, starting from the fundamental biology of CD70 and integrating the latest research advances with the current state of domestic development.
CD70, also known as TNFSF7, CD27L, or CD27LG, is part of the Tumor Necrosis Factor Receptor Super Family, which includes various protein ligands such as TNFa, lymphotoxin, CD30L, CD40L, FasL, OX40L, 4-1BBL, RANKL, APRIL, BLyS, NGF, and TRAIL. The human CD70 gene is located on chromosome 19p13 and codes for a 193-amino-acid trimer with a molecular weight of approximately 50 kDa. Structurally, CD70 consists of an extracellular binding region, a transmembrane segment, a short intracellular sequence, and two N-terminal glycosylated domains. The extracellular binding region shares similarities with other TNF ligand superfamily members. Disulfide bonds maintain CD70's specific spatial structure (Figure. 1) [1-5].
CD70 is naturally found on human and mouse medullary thymic epithelial cells (MTECs) as well as specific unconventional antigen-presenting cell (APC) subpopulations in the intestinal lamina propria. However, its presence is transient on activated T cells, B cells, and dendritic cells, with tight regulation dictated by pattern recognition receptors (PRRs), T and B cell antigen receptors, and cytokines such as IL-1a, IL-12, TNF-a, and prostaglandin E2. Additionally, CD70 expression is influenced by co-stimulation effects from CD28 and CD40 [1-3, 6]. In instances of malignant transformation, epithelial cells and other cell types may also exhibit CD70 expression.
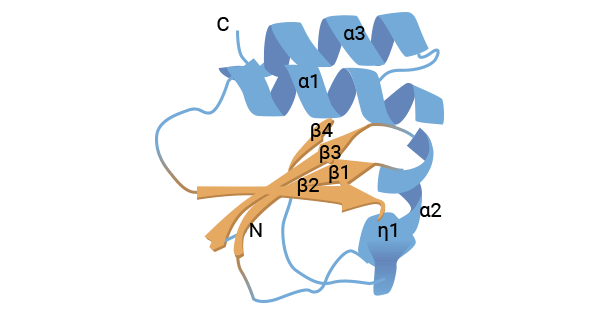
Figure 1. The structure of CD70 [5]
CD70 predominantly binds to its sole receptor, CD27, which belongs to the tumor necrosis factor receptor (TNFR) family alongside CD30, CD40, CD134 (OX40), and CD137 (4-1BB). These receptors play pivotal roles in regulating cell growth, differentiation, apoptosis, and programmed death. The CD27-CD70 interaction primarily delivers co-stimulatory signals crucial for the activation, proliferation, survival, and memory cell formation of T and B cells [6-8].
Various studies have revealed that CD27-CD70 co-stimulation supports the survival of developing Treg cells in the thymus, increases Treg numbers in the spleen and lymph nodes of tumor-bearing mice, and facilitates T-cell differentiation. Conversely, the absence of CD27-CD70 signaling impedes collateral circulation development in a mouse model of hindlimb ischemia and has implications in CD4+ T cell-mediated autoimmune diseases. Thus, the CD27-CD70 interaction is indispensable for regulating cellular immune responses (Figure 2) [9-11].
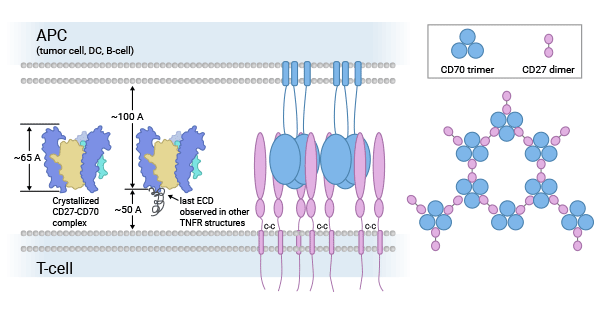
Figure 2. CD70-CD27 biological functions [7]
CD70/CD27L/TNFSF7 ligands activate CD27, leading to diverse biological responses. CD27 interacts with TRAFs like TRAF2 and TRAF5, triggering NF-kB and c-Jun kinase pathways to regulate cell proliferation, survival, and differentiation. It also binds to Siva 1, expanding its signaling functions. CD27's activation of the NF-kB pathway via NIK contributes to anti-apoptotic effects and complex immune modulation. Additionally, CD70 signaling activates PI3 and MAP kinase pathways, influencing cell cycle regulation in primary B cells and inducing cytotoxicity in various immune cells (Figure 3) [12-13].
Multiple animal studies have shown the vital role of the CD70-CD27 pathway in regulating T lymphocyte function. It's essential for the survival of virus-specific CD8+ T lymphocytes and contributes to the secretion of CXCL10 by activated CD8+T cells, suggesting a complex regulatory function among immune cells. CD27 signaling also enhances the differentiation of naïve CD4+ T cells into IFN-γ-secreting Th1 cells by promoting the Th1-specific transcription factor T-bet. Recent clinical trials in melanoma T-cell transfer have revealed a strong correlation between CD27 and CD70 expression on infused CD8+ tumor-infiltrating lymphocytes and antitumor effects, indicating the pathway's potential importance in cancer immunotherapy and providing insights into immune regulation mechanisms [14-16].
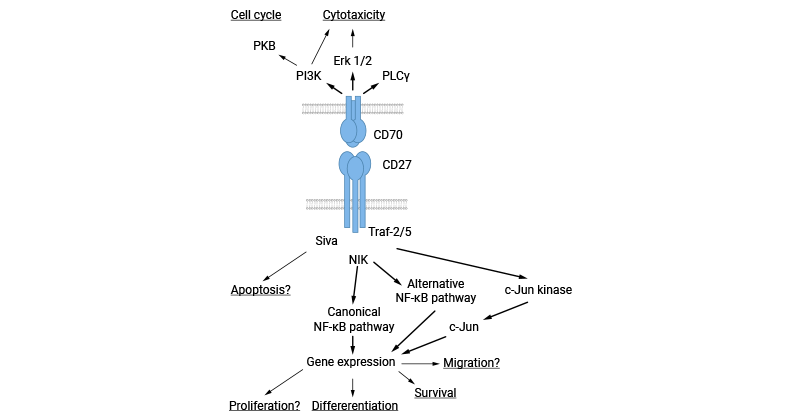
Figure 3. CD70-related signaling pathways [12]
CD70 serves as a crucial B-cell co-stimulatory molecule, fostering B-T cell interactions that drive B cell differentiation into plasma cells. This process boosts immunoglobulin production and enhances the cytotoxic effects of activated natural killer cells. Numerous pieces of evidence underscore CD70's pivotal role in various tumor types (e.g., lymphoma, solid tumors) and autoimmune diseases (e.g., lupus erythematosus). The following section summarizes studies exploring CD70's involvement in tumors and autoimmune conditions.
In hematologic malignancies, CD70 is commonly expressed in Hodgkin's lymphoma and lymphomas from B or T cells. It's also found in other B-cell-related cancers like diffuse large B-cell lymphoma, follicular lymphoma, B-lymphoblastic leukemia, multiple myeloma, and Waldenström's macroglobulinemia. In childhood acute lymphoblastic leukemia (ALL), CD27 and CD70 levels are significantly elevated in bone marrow, impacting leukemia cell proliferation. Blocking CD27-CD70 interaction with anti-human CD70 antibodies reduces leukemia cell growth in vitro. Though CD70's exact role in hematologic tumors isn't fully understood, its presence on tumor cells may reflect normal carcinogenesis. Targeting CD70/CD27 signaling shows promise for further research in treating AML [17-19].
CD70's widespread presence in solid tumors, coupled with its limited expression on normal cells, positions it as a promising target for novel antibody therapies against solid tumors. SGN-75, an antibody-drug conjugate (ADC), combines an anti-CD70 monoclonal antibody with the cytotoxic drug auristatin, offering versatile clinical applications. Auristatin, a potent cytotoxic agent, disrupts microtubule protein polymerization by binding to alpha-tubulin. The cytotoxic action of anti-CD70 ADCs involves intracellular internalization via the endosomal pathway, disrupting cellular microtubule networks, and arresting the cell cycle at the G2-M phase. Consequently, ADCs targeting CD70 selectively recognize and internalize tumor cells, delivering the drug to specific subcellular compartments for targeted tumor cell destruction [20-22].
CD70, a B-cell co-stimulatory molecule, is often overexpressed in systemic lupus erythematosus (SLE) patients, correlating with abnormal B-cell secretory processes. Inhibiting CD70 with specific antibodies can disrupt these aberrant processes. Various DNA methylation inhibitors, including Dnmt and ERK pathway inhibitors, have been found to elevate CD70 expression on peripheral blood CD4+ T cells. In patients with active lupus erythematosus, impaired ERK signaling, reduced Dnmt expression, and DNA hypomethylation contribute to CD70 overexpression on these T cells. Additionally, RFX1 recruits histone methyltransferase SUV39H1 to regulate CD70 expression, further suggesting the potential of CD70 antibodies as a treatment strategy for SLE [23-25].
The pathogenesis of rheumatoid arthritis involves immune system dysregulation, characterized by CD4+ T cell infiltration into synovial joints and the presence of MHC-I-positive antigen-presenting cells. B cells differentiate into plasma cells, producing immunoglobulins that form immune complexes and activate complement, triggering inflammation. In patients with rheumatoid arthritis, CD70 is expressed on the surface of CD4+ T cells, and its overexpression may result from aberrant regulation. This dysregulation can lead to the loss of CD28 expression, causing persistent surface CD70 expression after T cell activation [28-29].
Systemic scleroderma is a complex autoimmune disease characterized by immune dysfunction, vascular abnormalities, and fibrosis of organs' connective tissues. While its exact cause remains unclear, it's believed to result from a combination of genetic predisposition, environmental factors, and immune system abnormalities. Autoantibodies linked to systemic scleroderma, such as anti-nodule, anti-S170, anti-topoisomerase I, and anti-U1-RNP, are produced due to immune system dysregulation. Studies have revealed that in systemic scleroderma patients, the overexpression of CD70 on CD4+ T cells' surface correlates with hypomethylation of its regulatory sequences, suggesting a potential role for CD70 in the disease's pathogenesis [28-29].
Multiple sclerosis (MS) is an immune-related inflammatory disease affecting the central nervous system, particularly in females. While its exact cause is not fully understood, viral infections and immune abnormalities are implicated. Studies using experimental autoimmune encephalomyelitis, a model for MS, highlight the importance of CD70-CD27 interactions in T and B cell communication, crucial for antibody production. Blocking CD70 with anti-CD70 antibodies effectively inhibits experimental autoimmune encephalomyelitis by regulating TNF-a production, suggesting CD70 as a potential target for antibody-based therapy in autoimmune diseases [30].
In neurologic autoimmune diseases, CD70 has a dual role. Blocking the CD27-CD70 signaling pathway can inhibit autoimmune encephalomyelitis development. However, increased CD70 expression on certain B lymphocytes may worsen susceptibility to autoimmune encephalomyelitis. This highlights the complex interplay of CD70 activation in neuroinflammatory processes, where its timing and context can either promote or inhibit disease progression. Understanding these dynamics is vital for developing targeted therapies for neurologic autoimmune diseases [31].
Large-scale gene sequencing studies have linked the CD70 ligand CD27 gene to susceptibility to psoriasis. These studies also revealed reduced CD27 expression on T lymphocytes in psoriatic lesions and decreased plasma levels of soluble CD27 detected through immunohistochemical methods. While these findings offer valuable insights, further research is needed to elucidate the precise role of the CD27-CD70 pathway in psoriasis [33].
Moreover, CD70 has been implicated in inflammatory bowel disease (IBD). It is constitutively expressed on antigen-presenting cells (APCs) in the intestinal lamina propria, playing a crucial role in intestinal immune responses and homeostasis maintenance [34]. Recent studies suggest that CD70-expressing T cells contribute to more severe IBD and graft-versus-host disease, producing higher levels of inflammatory factors. Interestingly, IFN-γ induces T cells to express CD70, thereby regulating inflammatory T-cell responses by modulating T-cell proliferation inhibition [35].
Currently, there are 53 CD70-based drugs in development, spanning a diverse array of types including monoclonal antibodies, CAR-T cells, ADCs, CAR-NK cells, CAR-NKT cells, fusion proteins, dendritic cell vaccines, and RNA vaccines. For example, Monoclonal antibodies modulate or inhibit CD70, exerting interventional effects on tumor cell growth and metastasis. ADCs deliver toxicity to CD70-positive tumor cells while also targeting additional markers such as CD30 and Tubulin. Conversely, CAR-T cells recognize and eliminate CD70-positive tumor cells via modified T cells, concurrently targeting antigens like BCMA and CD19.
CAR-NK cells employ modified NK cells to recognize and attack CD70-positive tumor cells, alongside targets such as IL15, IL15R, TGFβ, and CD19. Fusion proteins regulate or inhibit CD27 through protein binding, targeting markers like PDL1, CD70, CD47, and SIRPα. Dendritic cell vaccines activate the immune system against tumor cells, targeting CD70, CD19, CD40L, and others. RNA vaccines induce tumor cell expression of CD70, CD40L, and TLR4 fusion proteins, directing immune responses. All in all, CD70 presents extensive prospects in tumor and immune system disease treatment, with ongoing research and cooperation anticipated to yield innovative research strategies in this field.
CD70, a significant protein in immunology, holds promise across various disease research like lupus and rheumatoid arthritis underscores its therapeutic potential. Furthermore, in cancer treatment, CD70-based drugs, including monoclonal antibodies, CAR-T cells, and fusion proteins, demonstrate efficacy against hematological malignancies and solid tumors. Collaborative efforts between international pharmaceutical companies, domestic firms, and academic institutions drive the development of these treatments. As research progresses, CD70's multifaceted role continues to inspire innovative therapeutic strategies, offering hope for improved outcomes in cancer and autoimmune research.
In order to assist pharmaceutical companies in the clinical research of CD70 in hematologic and solid tumors, as well as autoimmune diseases, such as lupus erythematosus, rheumatoid arthritis, systemic scleroderma, multiple sclerosis, etc., CUSABIO is launching the CD70 active protein product to help you explore the mechanism of CD70 or its potential clinical value.
CUSABIO Protein CD70
Recombinant Human CD70 antigen (CD70), partial (Active) Code: CSB-MP004954HU1
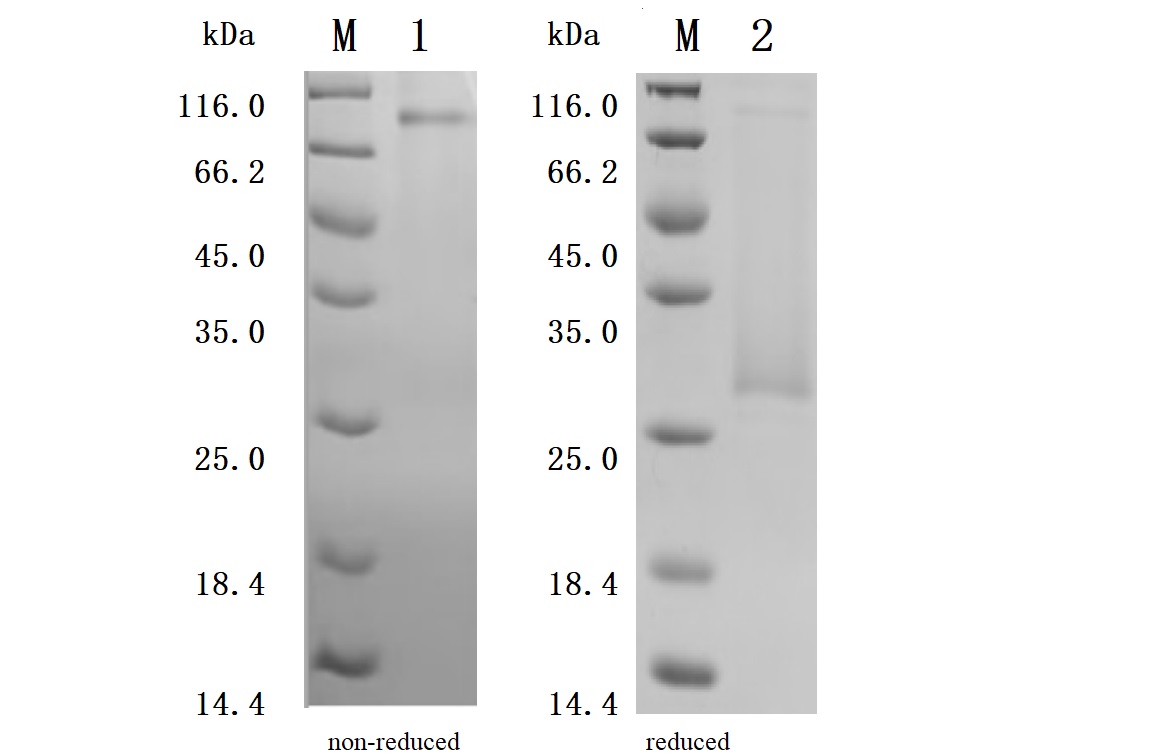
Purity was greater than 92.5% as determined by SDS-PAGE.
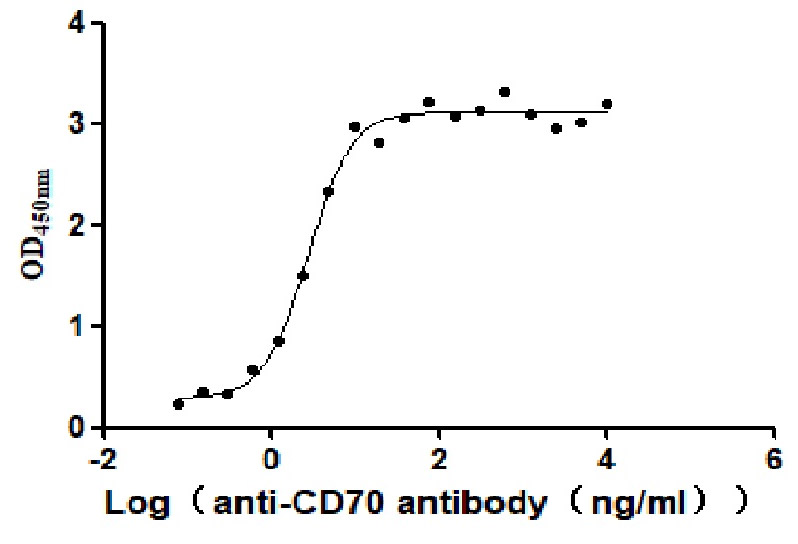
Immobilized Human CD70 at 2 μg/ml can bind Anti-CD70 antibody. The EC50 is 2.414-3.196 ng/mL.
References
[1] Aggarwal, Bharat B., et al. "The role of TNF and its family members in inflammation and cancer: lessons from gene deletion." Current Drug Targets-Inflammation & Allergy 1.4 (2002): 327-341.
[2] Watts, Tania H. "TNF/TNFR family members in costimulation of T cell responses." Annu. Rev. Immunol. 23 (2005): 23-68.
[3] Siegmund, Daniela, and Harald Wajant. "TNF and TNF receptors as therapeutic targets for rheumatic diseases and beyond." Nature Reviews Rheumatology 19.9 (2023): 576-591.
[4] Lubrano di Ricco, Martina, et al. "Tumor necrosis factor receptor family costimulation increases regulatory T‐cell activation and function via NF‐κB." European journal of immunology 50.7 (2020): 972-985.
[5] Dunne, Matthew, et al. "The CD27L and CTP1L endolysins targeting Clostridia contain a built-in trigger and release factor." PLoS pathogens 10.7 (2014): e1004228.
[6] Arroyo Hornero, Rebeca, et al. "CD70 expression determines the therapeutic efficacy of expanded human regulatory T cells." Communications biology 3.1 (2020): 375.
[7] Liu, Weifeng, et al. "Structural delineation and phase-dependent activation of the costimulatory CD27: CD70 complex." Journal of Biological Chemistry 297.4 (2021).
[8] Pandey, Arvind K., et al. "Expression of CD70 modulates nitric oxide and redox status in endothelial cells." Arteriosclerosis, Thrombosis, and Vascular Biology 42.9 (2022): 1169-1185.
[9] Lutfi, Forat, et al. "Targeting the CD27-CD70 pathway to improve outcomes in both checkpoint immunotherapy and allogeneic hematopoietic cell transplantation." Frontiers in Immunology 12 (2021): 715909.
[10] Flieswasser, Tal, et al. "The CD70-CD27 axis in oncology: the new kids on the block." Journal of experimental & clinical cancer research 41.1 (2022): 1-15.
[11] Abolhassani, Hassan. "Specific immune response and cytokine production in CD70 deficiency." Frontiers in Pediatrics 9 (2021): 615724.
[12] Borst, Jannie, Jenny Hendriks, and Yanling Xiao. "CD27 and CD70 in T cell and B cell activation." Current opinion in immunology 17.3 (2005): 275-281.
[13] Denoeud, Julie, and Muriel Moser. "Role of CD27/CD70 pathway of activation in immunity and tolerance." Journal of leukocyte biology 89.2 (2011): 195-203.
[14] Van De Ven, Koen, and Jannie Borst. "Targeting the T-cell co-stimulatory CD27/CD70 pathway in cancer immunotherapy: rationale and potential." Immunotherapy 7.6 (2015): 655-667.
[15] Peperzak, Victor, et al. "CD8+ T cells produce the chemokine CXCL10 in response to CD27/CD70 costimulation to promote generation of the CD8+ effector T cell pool." The Journal of Immunology 191.6 (2013): 3025-3036.
[16] Bowakim-Anta, Natalia, et al. "Chronic CD27-CD70 costimulation promotes type 1-specific polarization of effector Tregs." Frontiers in immunology 14 (2023): 1023064.
[17] Grewal, Iqbal S. "CD70 as a therapeutic target in human malignancies." Expert opinion on therapeutic targets 12.3 (2008): 341-351.
[18] Troeger, A., et al. "Significantly increased CD70 up regulation on TEL-AML positive B cell precursor acute lymphoblastic leukemia cells following CD40 stimulation." Klinische Pädiatrie (2014): 332-337.
[19] Sun, Qian, et al. "P317: ANTI-LEUKEMIA ACTIVITY OF CD70-DIRECTED IMMUNOTHERAPY IN B CELL PRECURSOR ACUTE LYMPHOBLASTIC LEUKEMIA." HemaSphere 7.S3 (2023): e6339407.
[20] Yang, Meijia, et al. "Tandem CAR-T cells targeting CD70 and B7-H3 exhibit potent preclinical activity against multiple solid tumors." Theranostics 10.17 (2020): 7622.
[21] Ryan, M. C., et al. "Targeting pancreatic and ovarian carcinomas using the auristatin-based anti-CD70 antibody–drug conjugate SGN-75." British journal of cancer 103.5 (2010): 676-684.
[22] Barreca, Marilia, et al. "An overview on anti-tubulin agents for the treatment of lymphoma patients." Pharmacology & therapeutics 211 (2020): 107552.
[23] Zhou, Y., et al. "Histone modifications and methyl-CpG-binding domain protein levels at the TNFSF7 (CD70) promoter in SLE CD4+ T cells." Lupus 20.13 (2011): 1365-1371.
[24] Balada, Eva, et al. "Associations between the expression of epigenetically regulated genes and the expression of DNMTs and MBDs in systemic lupus erythematosus." (2012): e45897.
[25] Zhao, Ming, et al. "RFX1 regulates CD70 and CD11a expression in lupus T cells by recruiting the histone methyltransferase SUV39H1." Arthritis research & therapy 12.6 (2010): 1-8.
[26] Nolte, Martijn A., et al. "Timing and tuning of CD27–CD70 interactions: the impact of signal strength in setting the balance between adaptive responses and immunopathology." Immunological reviews 229.1 (2009): 216-231.
[27] O’Neill, Rachel. Essential role of CD27/CD70 pathway in inflammatory T cell response. Diss. faculty of the Graduate School of the University at Buffalo, The State University of New York, 2019.
[28] Jiang, HongYan, et al. "Demethylation of TNFSF7 contributes to CD70 overexpression in CD4+ T cells from patients with systemic sclerosis." Clinical immunology 143.1 (2012): 39-44.
[29] Boutel, Maria Angeliki. Faculty Of Health Sciences. Diss. Aristotle University Of Thessaloniki, 2023.
[30] Dhaeze, Tessa, et al. "CD70 defines a subset of proinflammatory and CNS-pathogenic TH1/TH17 lymphocytes and is overexpressed in multiple sclerosis." Cellular & molecular immunology 16.7 (2019): 652-665.
[31] Lee, Eun Mi, et al. "CD70–CD27 ligation between neural stem cells and CD4+ T cells induces Fas–FasL-mediated T-cell death." Stem cell research & therapy 4.3 (2013): 1-10.
[32] Miller, James, et al. "Soluble CD70: a novel immunotherapeutic agent for experimental glioblastoma." Journal of neurosurgery 113.2 (2010): 280-285.
[33] Boursalian, Tamar E., et al. "Targeting CD70 for human therapeutic use." Therapeutic Targets of the TNF Superfamily (2009): 108-119.
[34] Manocha, Monika, et al. "Blocking CD27-CD70 costimulatory pathway suppresses experimental colitis." The Journal of immunology 183.1 (2009): 270-276.
[35] Leigh, Nicholas D., et al. "Host-derived CD70 suppresses murine graft-versus-host disease by limiting donor T cell expansion and effector function." The Journal of Immunology 199.1 (2017): 336-347.


Comments
Leave a Comment