Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Macrophages, central regulators of innate immunity and tissue homeostasis, exhibit substantial functional plasticity, with the most well-characterized dichotomy being the M1 and M2 subsets. This polarization balance is a fundamental hallmark of macrophage biology and a critical determinant of disease pathogenesis, therapeutic efficacy, and tissue regeneration, making it a focus of biomedical research. Elucidating M1/M2 characteristics is crucial for understanding immune-related disorder mechanisms, developing targeted therapies, and advancing immunological research.
This article systematically describes the M1/M2 macrophage phenotypic, molecular, and functional properties, along with pathophysiological roles and research methods, to inform basic and translational investigations.
Table of Contents
2. Molecular Drivers and Signaling Pathways that Affect Macrophage Polarization
3. Research Methods and Technical Advances in Macrophage Phenotypic Identification
4. Macrophages' Functional Roles in Disease Pathogenesis and Resolution
Macrophages aren't rigidly programmed cells—instead, they have remarkable plasticity, allowing them to adjust their functions in response to local environmental signals. This functional specialization is known as polarization. Macrophages are derived from circulating monocytes or tissue-resident progenitors and are primarily divided into two archetypes: the pro-inflammatory M1 and the reparative M2 macrophages.
The M1/M2 binary model was proposed in the early 2000s, drawing inspiration from the Th1 and Th2 classification of T helper cells. M1 macrophages, often classically activated by signals like IFN-γ and lipopolysaccharide (LPS), are the body's first line of defense against pathogens and tumors. They produce high levels of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6, generate reactive nitrogen and oxygen species (ROS), and promote Th1 immune responses [1,2]. They primarily orchestrate a robust inflammatory response to eliminate pathogens and damaged cells.
In contrast, M2 macrophages, alternatively activated by cytokines like IL-4 and IL-13, are involved in resolving inflammation, clearing cellular debris, and promoting tissue remodeling and repair, angiogenesis, and immunoregulation [1]. They express markers like CD206 and ARG-1 and secrete anti-inflammatory cytokines such as IL-10 and TGF-β [3,4]. The M2 phenotype can be further subdivided into: M2a, M2b, M2c, and M2d.
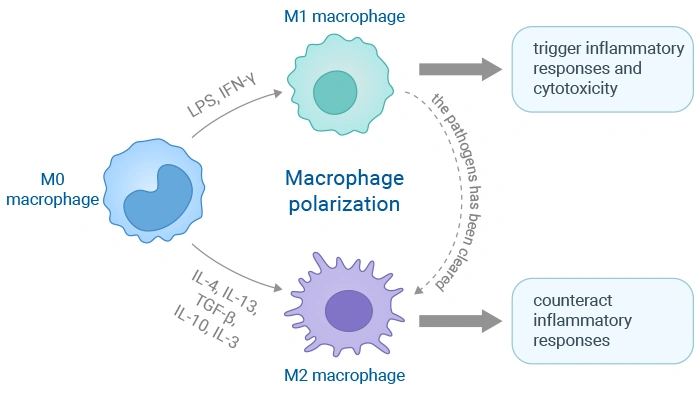
Figure 1. The polarization process of macrophages and its triggering targets
Table 1. Phenotypes, stimulants, biomarkers, secretions, and functions of macrophages [12].
| Macrophage phenotype | Stimulants | Stimulants | Secretions | Functions |
|---|---|---|---|---|
| M1 (classically activated macrophages) | IFN-γ, LPS, TNF-α, GM-CSF | CD86, CD40, CD38, NF-κB, STAT1 | TNF-α, IL-1α, IL-12, IL-23, IL-1β, IL-6, ROS, and RNS | Promote Th1 immune response, promote inflammatory response, fight pathogens, and inhibit the occurrence and development of tumors |
| M2a (wound healing macrophages) | IL-4, IL-13, M-CSF | CD206, IL-1R, CCL17, Fizz1, STAT6 | TGF-β, IL-10, insulin-like growth factor (IGF), and fibronectin | Promote tissue repair and remodeling, promote fibrosis, and promote type II immune response by enhancing polyamines, collagen synthesis |
| M2b (regulatory macrophages) | Immune complex, TLR agonist, IL-1R agonist | IL-10, CCL1, LIGHT, CD86, SPHK1, TNF-α, IL-6, ERK, AP-1 | Proinflamatory cytokines (IL-1β, IL-6, and TNF-α), anti-inflammatory cytokine (IL-10 and low levels of IL-12) | Involve in proinflammatory and anti-inflammatory responses, immunomodulation, and Th2 activation |
| M2c (acquired inactivated macrophages) | Glucocorticoids, TGF-β, IL-10 | CD163, MerTK, STAT3 | IL-10, TGF-β | Immune tolerance and tissue repair, suppress inflammation, promote phagocytosis and cholesterol efflux |
| M2d (tumor-associated macrophages) | TLR, adenosine A2A receptor γ, IL-6 |
VEGF-A, HIF-1α | Proteolytic enzymes (MMP-2), growth factors (VEGF), and anti-inflammatory mediators (TGF-β) | Beneficial for angiogenesis and tumor metastasis |
Macrophage polarization into classically activated (M1) or alternatively activated (M2) phenotypes is a fundamental process in immunity, inflammation, and tissue repair. This functional switch is tightly regulated by a complex network of intracellular signaling pathways [5,6], which respond to specific microenvironmental cues.
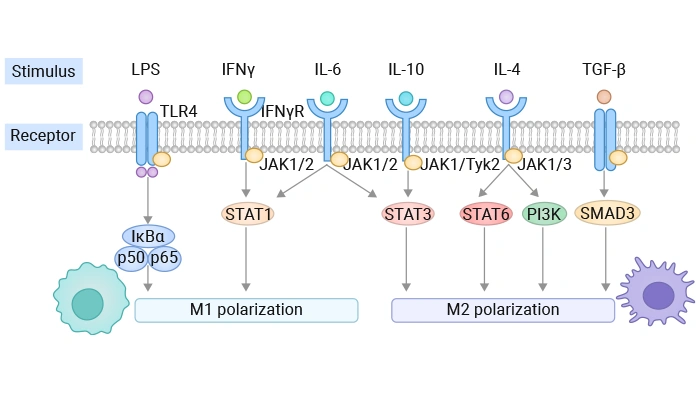
Figure 2. Factors affecting macrophage polarization
Research on macrophage phenotypes has moved well beyond simple M1/M2 marker panels and now integrates multi‑omic, imaging, and biophysical approaches for more precise identification.
Table 2. A summary of the macrophage phenotyping methods.
| Method Category | Specific Method | Key Principles / Measured Parameters | Key Features / Applications |
|---|---|---|---|
| Classical Methods | Flow Cytometry & Immunophenotyping | Panels of surface/intracellular markers (e.g., CD64, CD86 for M1; CD206, Arg1 for M2). | Uses polychromatic panels (e.g., 11-color) with partially redundant markers for robust classification [13,14]; species-adapted panels validated via transcriptomics and functional assays [15]. |
| Gene Expression Assays (RT-qPCR & Bulk Transcriptomics) | RT‑qPCR is widely used to quantify hallmark cytokines and polarization markers (IL1B, IL6, IL10, NOS2, ARG1, etc.) and to validate M1/M2 states alongside protein‑level readouts [16]. | Bulk RNA‑seq and microarray studies enabled early “spectrum” models of macrophage activation, revealing that polarization in response to diverse stimuli occupies a continuum rather than discrete M1/M2 states; network-based analyses of bulk transcriptomes have identified gene modules associated with specific activation cues, providing a framework for more nuanced classification [17]. | |
| Secretome & Protein-based Assays (ELISA, Multiplex Bead Arrays, WB) | ELISA and multiplex bead arrays are routinely used to profile cytokines and chemokines (e.g., TNF, IL‑1β, IL‑6, IL‑10, CXCL10) secreted by polarized macrophages; WB and immunoblot-based workflows are used to validate newly identified protein biomarkers, such as polarization-associated membrane proteins discovered by proteomics. | Complementing flow and qPCR data; essential for phenotype verification in human and animal models [18]. | |
| Advanced Imaging & Biophysical Approaches | Morphology-based High-Content Imaging | Image‑based machine learning analysis of morphometric features (size, roundness, elongation, protrusions) [19]. | Classifies M0, M1, M2 subtypes (and even M2a vs M2c subtypes) with ~93% accuracy; links morphology to functional output (e.g., IL-10 prediction). |
| Membrane-Order-Sensitive Probes (e.g., Di-4-ANEPPDHQ) | A 2025 study compared RT‑qPCR, flow cytometry, and the membrane-order probe Di‑4‑ANEPPDHQ in THP‑1-derived macrophages, showing that the probe distinguishes M0, M1, and M2 phenotypes via distinct red/blue fluorescence shifts corresponding to depolarized (M1) and hyperpolarized (M2) membranes [16]. | Integrating Di‑4‑ANEPPDHQ imaging with gene and surface marker analysis improved sensitivity and provided real‑time information on dynamic changes in macrophage activation state. | |
| Live-Cell Imaging & Functional Readouts | Time‑lapse microscopy, often combined with fluorescent reporters, allows direct visualization of phagocytosis, migration, and interaction with other cells [19]. | Links phenotypic state to dynamic functional behavior; combined with single-cell tracking and machine learning to connect behavioral patterns with molecular signatures. | |
| Single-Cell & Spatial Omics | Single-Cell RNA Sequencing (scRNA-seq) | Unsupervised identification of transcriptionally distinct macrophage subsets within tissues. | Reveals population heterogeneity beyond M1/M2 (e.g., profibrotic subsets) [19,20]; defines novel subpopulations. |
| Spatial Transcriptomics & Multiplex Imaging | Overlays gene or protein expression data onto tissue architecture to localize macrophage subsets within specific niches, such as fibrotic regions or tumor invasive fronts. | Distinguishes resident vs. infiltrating cells in situ combined with multiplex immunofluorescence or imaging mass cytometry; crucial for understanding how local microenvironments shape macrophage states in fibrosis, cancer, and infection [19,20] | |
| Multi-Omics Integration | Integrates scRNA‑seq with surface proteomics (CITE‑seq), chromatin accessibility (scATAC‑seq), and metabolomic signatures to build multi‑layered maps of macrophage states [17,19]. | Better capture activation networks and lineage relationships; supports data-driven phenotypic definitions. | |
| Functional Assays | Antimicrobial & Phagocytic Assays | Phagocytosis assays using fluorescent beads, bacteria, or apoptotic cells are used to quantify uptake capacity across phenotypes [15]; antimicrobial functional assays (bacterial killing or parasite control readouts) have been combined with transcriptomics and flow cytometry to correlate effector functions with surface marker and gene expression profiles in species such as dog and mouse [19]. | Correlates effector functions with molecular signatures from omics or flow cytometry data. |
| Cytokine Production & Signaling Dynamics | Measures stimulus-induced cytokine release by ELISA or multiplex assays, combined with phospho‑flow cytometry for signaling intermediates (e.g., STAT1, STAT3, NF‑κB) [17,19]. | Provides a dynamic perspective on responsiveness; helps distinguish transient versus stable activation states. | |
| Emerging & Integrative Approaches | Machine Learning & Computational Phenotyping | Algorithms integrating multi-dimensional data (morphology, flow, transcriptomics) for classification [17,19]. | Enhances classification accuracy beyond single-modality approaches; can predict functional outputs like cytokine levels [14]. |
| Biophysical & Microenvironment-Mimicking Platforms | Microfluidic and biophysical platforms are being developed to study macrophages under controlled shear stress, stiffness, and chemokine gradients, enabling more physiologic phenotyping [19]. | Organoid and organ‑on‑chip systems that co‑culture macrophages with tissue‑specific cells (e.g., epithelium, endothelium, tumor cells) allow to examine how complex microenvironments drive context-specific phenotypes beyond simple M1/M2 states. |
Macrophages are central orchestrators of the immune response, playing a dual and paradoxical role in human health and disease. Their activities span from initiating and sustaining inflammation to actively promoting its resolution and facilitating tissue repair.
Tumor-associated macrophages (TAMs) often exhibit an M2-like, pro-tumorigenic phenotype, supporting immunosuppression, angiogenesis, and metastasis [21-23]. However, evidence shows cancers like triple-negative breast cancer can induce a mixed M1/M2 phenotype that still promotes aggressiveness [24]. Furthermore, in certain contexts like oral squamous cell carcinoma, M1-like TAMs may also contribute to tumor progression [25]. The therapeutic goal is often to repolarize TAMs from M2 to M1 to reinstate anti-tumor immunity [11,23].
In rheumatoid arthritis (RA), synovitis is characterized by a dominant M1/Th1 response that drives joint destruction via pro-inflammatory cytokines and matrix metalloproteinases [2]. Shifting the balance toward M2/Th2 can promote remission. Similarly, in atherosclerosis, M1 macrophages contribute to plaque instability, while M2 macrophages promote resolution [26,27]. Agents like ELABELA show therapeutic potential by restoring this balance [27].
The timely transition from M1 to M2 is critical for wound healing. However, persistent M2 activation or the emergence of specific profibrotic subsets, such as CD9+TREM2+ macrophages driven by type 3 inflammation (GM-CSF, IL-17A), can lead to pathological fibrosis in organs like the liver and lung [28]. Conversely, in necrotizing enterocolitis (NEC), administering M2-derived nanovesicles to foster M2 polarization can resolve damaging intestinal inflammation [29].
In chronic liver diseases, omega-3 fatty acids and their specialized pro-resolving mediators (e.g., MaR1) protect by promoting a beneficial M2 polarization, reducing oxidative stress and inflammation [30]. In retinopathy, the natural progression involves a phase change where early M1 activity drives neovascularization, followed by a late M2 phase that promotes regression, suggesting timed therapeutic interventions [7].
This is where fundamental research translates into clinical potential. Detail the innovative approaches being developed.
Macrophages are versatile immune cells whose functional polarization into pro-inflammatory M1 or reparative M2 states is central to health and disease. This balance, driven by specific signaling pathways like JAK-STAT and NF-κB, dictates outcomes in cancer, chronic inflammation, and tissue repair. In pathologies, the equilibrium is often disrupted, for example, in tumors, where M2-like macrophages promote immunosuppression. Consequently, therapeutic strategies now aim to reprogram macrophage polarization using nanotechnology for targeted delivery, biological agents, and metabolic interventions. Looking ahead, the field is moving beyond the simple M1/M2 dichotomy. Future research will leverage single-cell analyses to decipher macrophage heterogeneity and develop context-dependent, precision therapies that dynamically modulate these critical cells to restore immune homeostasis and improve clinical outcomes.
References
[1] Yunna C, Mengru H, Lei W, Weidong C. Macrophage M1/M2 polarization [J]. Eur J Pharmacol. 2020 Jun 15;877:173090.
[2] Cutolo M, Campitiello R, Gotelli E, Soldano S. The Role of M1/M2 Macrophage Polarization in Rheumatoid Arthritis Synovitis [J]. Front Immunol. 2022 May 19;13:867260.
[3] Zhang X, Li J, Liu Y, et al. Myricetin alleviates DNCB-induced atopic dermatitis by modulating macrophage M1/M2 polarization [J]. Int Immunopharmacol. 2025 Oct 10;163:115212.
[4] Hung CH, Hsu HY, et al. Arsenic Induces M2 Macrophage Polarization and Shifts M1/M2 Cytokine Production via Mitophagy [J]. Int J Mol Sci. 2022 Nov 10;23(22):13879.
[5] Strizova, Z., Benesova, I., et al. (2023). M1/M2 macrophages and their overlaps – myth or reality [J]? Clinical Science (London, England: 1979), 137(15), 1067.
[6] Wang, N., Liang, H., & Zen, K. (2014). Molecular Mechanisms That Influence the Macrophage M1–M2 Polarization Balance [J]. Frontiers in Immunology, 5, 614.
[7] Li J, Yu S, et al. The phase changes of M1/M2 phenotype of microglia/macrophage following oxygen-induced retinopathy in mice [J]. Inflamm Res. 2021 Feb;70(2):183-192.
[8] Pagie, S., Gérard, N., & Charreau, B. (2018). Notch signaling triggered via the ligand DLL4 impedes M2 macrophage differentiation and promotes their apoptosis [J]. Cell Communication and Signaling: CCS, 16, 4.
[9] Miki, S., Suzuki, J. I., et al. (2021). S-1-Propenylcysteine promotes IL-10-induced M2c macrophage polarization through prolonged activation of IL-10R/STAT3 signaling [J]. Scientific Reports, 11, 22469.
[10] Sun X, Li Z, Wang X, He J, Wu Y. Inorganic Phosphate as "Bioenergetic Messenger" Triggers M2-Type Macrophage Polarization [J]. Adv Sci (Weinh). 2024 Apr;11(13):e2306062.
[11] Liu H, Yao M, Ren J. Codonopsis pilosula-derived glycopeptide dCP1 promotes the polarization of tumor-associated macrophage from M2-like to M1 phenotype [J]. Cancer Immunol Immunother. 2024 May 14;73(7):128.
[12] Peng, Y., Zhou, M., et al. (2023). Regulatory Mechanism of M1/M2 Macrophage Polarization in the Development of Autoimmune Diseases [J]. Mediators of Inflammation, 2023, 8821610.
[13] Liu L, Stokes JV, Tan W, Pruett SB. An optimized flow cytometry panel for classifying macrophage polarization [J]. J Immunol Methods. 2022 Dec;511:113378.
[14] Selig, M., Poehlman, L., Lang, N. C., Völker, M., Rolauffs, B., & Hart, M. L. (2024). Prediction of six macrophage phenotypes and their IL-10 content based on single-cell morphology using artificial intelligence [J]. Frontiers in Immunology, 14, 1336393.
[15] Chow, L., Soontararak, S., Wheat, W., Ammons, D., & Dow, S. (2022). Canine polarized macrophages express distinct functional and transcriptomic profiles [J]. Frontiers in Veterinary Science, 9, 988981.
[16] Siniyeh, A. A., Alshaer, W., et al. (2025). Comparative analysis of RT-qPCR, flow cytometry, and Di-4-ANEPPDHQ fluorescence for distinguishing macrophage phenotypes [J]. Biochemistry and Biophysics Reports, 44, 102225.
[17] Xue, J., Schmidt, S. V., et al. (2014). Transcriptome-Based Network Analysis Reveals a Spectrum Model of Human Macrophage Activation [J]. Immunity, 40(2), 274.
[18] Liu, B., Nguyen, P. L., Li, P., Naldrett, M. J., Alvarez, S., & Yu, J. (2025). Identification of novel protein biomarkers of macrophage polarization using comparative proteomic analyses of murine primary macrophages [J]. The Journal of Immunology Author Choice, 214(10), 2730.
[19] Vishnyakova, P., Elchaninov, A., Fatkhudinov, T., & Kolesov, D. (2025). Unravelling approaches to study macrophages: From classical to novel biophysical methodologies [J]. PeerJ, 13, e19039.
[20] Coulis, G., Jaime, D., et al. (2023). Single-cell and spatial transcriptomics identify a macrophage population associated with skeletal muscle fibrosis [J]. BioRxiv, 2023.04.18.537253.
[21] Li S, Fu X, Ning D, et al. Colon cancer exosome-associated HSP90B1 initiates pre-metastatic niche formation in the liver by polarizing M1 macrophage into M2 phenotype [J]. Biol Direct. 2025 Apr 15;20(1):52.
[22] Wang Q, Ma W. Revisiting TAM polarization: beyond M1- and M2-type TAM toward clinical precision in macrophage-targeted therapy [J]. Exp Mol Pathol. 2025 Sep;143:104982.
[23] Shan H, Dou W, Zhang Y, Qi M. Targeted ferritin nanoparticle encapsulating CpG oligodeoxynucleotides induces tumor-associated macrophage M2 phenotype polarization into M1 phenotype and inhibits tumor growth [J]. Nanoscale. 2020 Nov 12;12(43):22268-22280.
[24] Pe KCS, Saetung R, et al. Triple-negative breast cancer influences a mixed M1/M2 macrophage phenotype associated with tumor aggressiveness [J]. PLoS One. 2022 Aug 12;17(8):e0273044.
[25] Li CX, Gong ZC, Yu JW. Deliberation concerning the role of M1-type macrophage subset in oral carcinogenesis [J]. J Exp Clin Cancer Res. 2024 Aug 8;43(1):220.
[26] Shou X, Wang Y, Jiang Q, Chen J, Liu Q. miR-126 promotes M1 to M2 macrophage phenotype switching via VEGFA and KLF4 [J]. PeerJ. 2023 Mar 31;11:e15180.
[27] Tang L, Yi X, Tan W, et al. ELABELA Ameliorates Atherosclerosis Through Restoring the M1/M2 Macrophage Balance in ApoE(-/-) Mice [J]. J Am Heart Assoc. 2025 Jul 15;14(14):e041261.
[28] Fabre T, Barron AMS, et al. Identification of a broadly fibrogenic macrophage subset induced by type 3 inflammation [J]. Sci Immunol. 2023 Apr 14;8(82):eadd8945.
[29] Liu, C., Gong, J., et al. (2025). M2-type macrophage nanovesicles regulate the inflammatory response after necrotizing enterocolitis by inducing M1 to M2-like macrophage polarization [J]. Frontiers in Cellular and Infection Microbiology, 15, 1664897.
[30] Videla LA, Valenzuela R, Del Campo A, Zúñiga-Hernández J. Omega-3 Lipid Mediators: Modulation of the M1/M2 Macrophage Phenotype and Its Protective Role in Chronic Liver Diseases [J]. Int J Mol Sci. 2023 Oct 24;24(21):15528.
[31] Zhang Y, Chu B, Fan Q, Song X, Xu Q, Qu Y. M2-type macrophage-targeted delivery of IKKβ siRNA induces M2-to-M1 repolarization for CNV gene therapy [J]. Nanomedicine. 2024 Apr;57:102740.
[32] Wang, P., Li, Z., Song, Y., Zhang, B., & Fan, C. (2025). Resveratrol-driven macrophage polarization: Unveiling mechanisms and therapeutic potential [J]. Frontiers in Pharmacology, 15, 1516609.


Comments
Leave a Comment