
Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-RA008790A0HU |
| Size | US$210 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
| Application | Recommended Dilution |
|---|---|
| WB | 1:500-1:5000 |
| IHC | 1:50-1:200 |
| IF | 1:20-1:200 |
FOS, also known as c-Fos, serves as a critical immediate-early gene product that functions as a key component of the AP-1 transcription factor complex. This proto-oncogene plays essential roles in signal transduction pathways linking extracellular stimuli to changes in gene expression, making it particularly valuable for neuroscience research where it serves as a widely-used marker of neuronal activation. Its involvement in cell proliferation, differentiation, and survival also positions FOS as an important target in cancer biology and cellular stress response studies.
This recombinant monoclonal antibody, clone 14C10, offers the reproducibility and consistency that demanding research applications require. Because recombinant antibodies are produced from defined sequences rather than traditional hybridoma methods, you can expect reliable performance across experiments and between lots, eliminating a common source of variability in long-term studies.
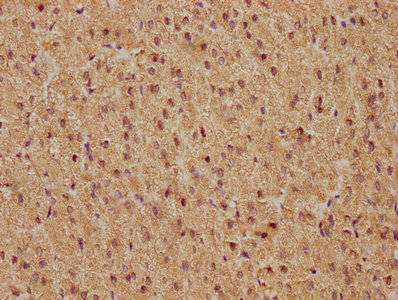
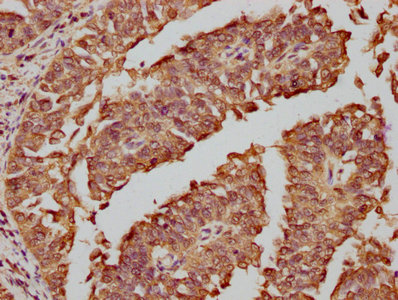
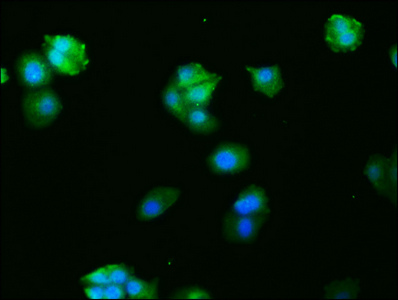
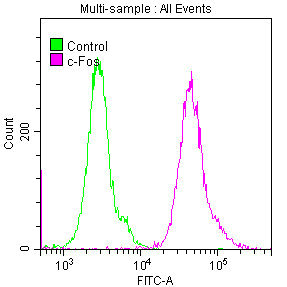
Validation data demonstrates robust performance across multiple experimental platforms. Western blot analysis confirms detection in both human cell lines including HepG2, HeLa, MCF-7, Jurkat, A549, and 293, as well as mouse NIH/3T3 cells, supporting the antibody's cross-species reactivity. The observed band at 62 kDa, higher than the predicted molecular weight, likely reflects post-translational modifications such as phosphorylation, which is characteristic of FOS regulation during cellular activation. Immunohistochemistry validation in human adrenal gland and cervical cancer tissues demonstrates suitability for tissue-based studies, while immunofluorescence staining in HepG2 cells and flow cytometry analysis in HeLa cells expand your options for cellular localization and quantitative single-cell analyses.
Whether you are mapping neuronal activity patterns, investigating oncogenic signaling, or studying immediate-early gene responses, this antibody provides the experimental flexibility and recombinant-derived consistency to support your research objectives.
There are currently no reviews for this product.

