Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)

Hepatitis viruses are a group of viruses that primarily affect the liver, including hepatitis A virus (HAV), hepatitis B virus (HBV), hepatitis C virus (HCV), hepatitis D virus (HDV), and hepatitis E virus (HEV). HAV and HEV are mainly associated with fecal–oral transmission, while HBV, HCV, and HDV are primarily transmitted through infected blood or body fluids. These viruses differ in genome type, infection outcome, life cycle, and viral protein composition. Among the five major hepatitis viruses, HBV is the only DNA virus, while HAV, HCV, HDV, and HEV are RNA viruses.
This resource provides an overview of the key characteristics, life cycles, viral proteins, and related research products for HAV, HBV, HCV, HDV, and HEV, supporting researchers in virology, infectious disease, liver disease, and host-virus interaction studies.
Table 1. Characteristics of hepatitis viruses
| HAV | HBV | HCV | HDV | HEV | |
|---|---|---|---|---|---|
| Family | Picornaviridae | Hepadnaviridae | Flaviviridae | Not applicable | Hepeviridae |
| Genus | Hepatovirus | Orthohepadnavirus | Hepacivirus | Deltavirus | Orthohepevirus |
| Genome | Positive-sense single-stranded linear RNA | Double-stranded DNA | Positive-sense single-stranded linear RNA | Negative-sense single-stranded circular RNA | Positive-sense single-stranded linear RNA |
| Transmission | Fecal–oral | Exposure to infected blood or body fluid/sexual/perinatal | Exposure to infected blood | Exposure to infected blood/body fluids | Fecal–oral/zoonotic/blood transfusion |
| Clinical outcome of infection | Self-limited | Self-limited and chronic | Self-limited and chronic | Self-limited and chronic | Self-limited |
The Table information is cited from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8999150/
HAV is a positive-sense single-stranded RNA virus mainly associated with fecal–oral transmission.
The HAV genome encodes a single polyprotein that is processed into structural capsid proteins and non-structural proteins involved in viral replication.
HAV-related research often focuses on viral entry, polyprotein processing, capsid proteins, and host-virus interaction.
HAV is a non-enveloped RNA virus with a 7.5 kb genome. Its genome encodes a single large polyprotein that is processed into four capsid proteins (VP1, VP2, VP3, VP4) and seven non-structural polypeptides (2A, 2B, 2C, 3A, 3B, 3C, 3D). Infectious HAV virions exist in two forms: naked, non-enveloped HAV (neHAV) virions released into the stool, and quasi-enveloped virions (eHAV) generated by hijacking control of the host.
| HAV protein | Function | ||
|---|---|---|---|
| Genomic polyprotein | Structural proteins | VP1 | Make up the capsid polypeptide |
| VP2 | |||
| VP3 | |||
| VP4 | |||
| Non-structural proteins | 2A | Required for viral replication | |
| 2B | |||
| 2C | |||
| 3A | Anchor the replication component to the cell membranes | ||
| 3B | Also call VPg | ||
| 3C | A cysteine protease that cleaves the polypeptides into proteins | ||
| 3D | An RNA polymerase | ||
HAV enters host hepatocytes by engaging with cell surface molecules, specifically sialic acid and ganglioside, followed by the uncoating of viral capsid and subsequent release of viral RNA from endosomes into the cytoplasm [1]. The viral genome is translated into a polyprotein under the regulation of an internal ribosome entrance site (IRES) and then cleaved into distinct structural and nonstructural proteins.
The genomic RNA is also used as a template to generate numerous new replicas of the RNA genome, which are packed into capsids to produce intracellular viral offspring. Newly synthesized HAV virions are released from cells in quasi-enveloped forms.
HBV is the only DNA virus among the five major hepatitis viruses and contains a partially double-stranded relaxed circular DNA genome.
Key HBV proteins include HBsAg, HBcAg, HBeAg, HBx, and DNA polymerase, which are involved in viral structure, replication, immune response, and host-virus interaction.
HBV research commonly focuses on viral entry, NTCP/SLC10A1-mediated infection, cccDNA formation, viral protein function, and chronic infection mechanisms.
The HBV virion consists of an outer lipoprotein envelope composed of hepatitis B surface antigen (HBsAg), and a hepatitis B core antigen (HBcAg)-formed icosahedral nucleocapsid which encapsulates a partially double-stranded, relaxed circular DNA (rcDNA) genome of 3.2 kb.
The HBV genome consists of four partially overlapping and open reading frames (ORFs), C (core), P (polymerase), X (regulatory X protein), and S (surface), from which seven functional proteins are synthesized: HBcAg, HBeAg, polymerase (pol), HBx, and three types of HBsAg - the small (s), medium (M), and large (L) proteins.
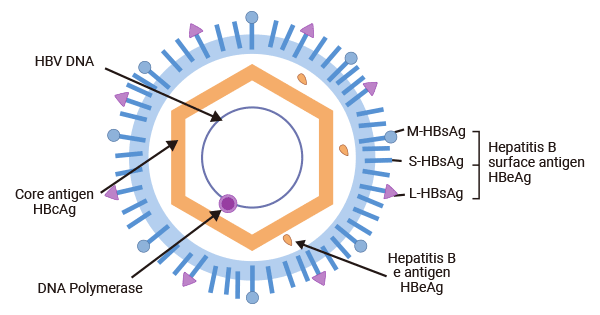
Figure 1. HBV structure
The picture is cited from Wikipedia
| HBV proteins | Function |
|---|---|
| HBcAg | Assemble intothe icosahedral viral nucleocapsid to enclose the HBV genome and DNA polymerase |
| HBeAg | Not necessary for viral replication or infection; an indicator of active viral replication and is linked to the establishment of chronic infection, hepatic inflammatory injury, and HCC development |
| DNA Polymerase | Initiate viral replication by reverse transcription |
| HBx | Localized in different subcellular compartments, conferring distinct functions; involved in various cellular events, including transactivator of the viral and cellular promoters, epigenetic modifications, ubiquitination, autophagy and non-coding RNA regulation, to promote hepatocellular carcinogenesis, stabilization and activation of cccDNA |
| HBsAg | Include L-HBsAg, M-HBsAg, and S-HBsAg; make up the viral envelope |
HBV enters into the host hepatocytes in an endocytosis-dependent manner. HBV initiates by binding to heparan sulfate proteoglycans (HSPGs) such as glypican 5. And then, it engages with the sodium taurocholate cotransporting peptide (NTCP/SLC10A1) receptor on the surface of hepatocytes to elicit viral internalization [2-4].
Studies have demonstrated that EGFR triggers HBV’s internalization into host cells by interacting directly with NTCP [5,6]. Following endocytosis, the viral nucleocapsid is released into the hepatic cytoplasm and then transported to the nucleus.
In the nucleus, the viral genomic rcDNA is converted to cccDNA (covalently closed circular DNA) to amplify a viral RNA intermediate, which is then reverse-transcribed back to viral DNA [7]. Simultaneously, the viral DNA integrates into the host genome, which is the cause that hepatitis B is incurable.
The cccDNA minichromosome remains in the nucleus, serving as a template for viral RNA transcription. It encodes seven essential viral proteins necessary for replication: three different sizes of HBsAg, HBcAg, HBeAg, HBx, and DNA polymerase [8].
In the cytoplasm, HBcAg proteins self-assemble into an icosahedral nucleocapsid, encapsulating viral polymerase and pgRNA. Following reverse transcription within the nucleocapsid, viral capsid that contains the HBV genome binds to HBsAg proteins in the endoplasmic reticulum for encapsulation. Ultimately, mature HBV virions are released from hepatocytes through multivesicular bodies (MVBs).
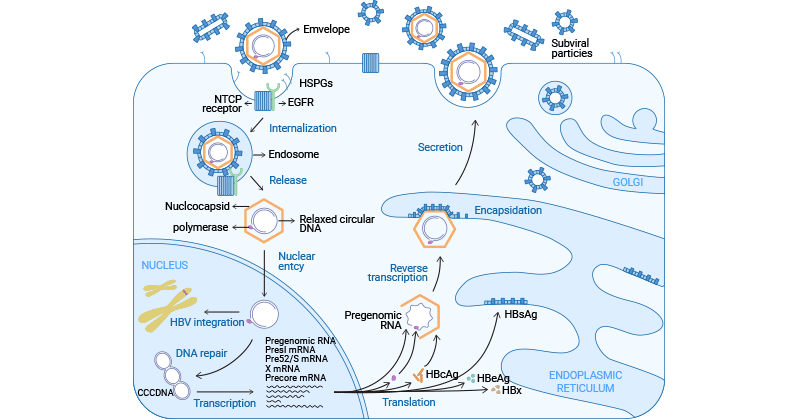
Figure 2. HBV life cycle
The picture is cited from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8387624/
HCV is a positive-sense single-stranded RNA virus with a genome that encodes one polyprotein precursor.
The HCV polyprotein is processed into structural proteins, including core, E1, and E2, and non-structural proteins such as p7, NS2, NS3, NS4A, NS4B, NS5A, and NS5B.
HCV-related studies often focus on viral entry, envelope protein function, RNA replication, polyprotein processing, and host-cell interaction.
HCV has an RNA genome of about 9.6 kb in length. The viral genome contains a single open reading frame (ORF) that encodes a single polyprotein precursor, which is subsequently processed into three structural proteins and seven non-structural proteins.
| HCV Proteins | Function | |||
|---|---|---|---|---|
| Genome polyprotein | Structural proteins | Capsid (C) (p22) | Make up the viral nucleocapsid | |
| Envelope glycoproteins | E1 (gp35) | Major components of the virion envelope; required for viral entry and fusion | ||
| E2 (gp70) | ||||
| Non-structural proteins | NS1 (P7) | Calcium ion channel necessary for the efficient recruitment and release of the proliferating virus | ||
| NS2 (p23) | autoprotease | |||
| NS3 (p70) | Serin protease/RNA helicase | |||
| NS4A (p8) | NS3 protease cofactor | |||
| NS4B (p27) | Formation of membranous web | |||
| NS5A (pp56/58) | Involvement in viral replication, packaging, and release of HCV particles | |||
| NS5B (p68) | RNA-dependent RNA-polymerase | |||
HCV enters the cell in a pH and clathrin-dependent endocytosis. HCV first attaches to the host cells by binding to HSPG and LDL-R. After docking, the HCV particle binds to CD81 and scavenger receptor class B type I (SR-BI) on the surface of the hepatocyte through E2 [9,10] and then interacts with claudin-1 and occludin, which facilitates its entry into the cell with the endosome [11].
Following endosome-mediated internalization, endosomal acidification promotes low pH-dependent HCV envelope fusion with the endosomal membrane, resulting in the release of viral genomic RNA into the cytosol [12]. The RNA is used as the template both for replication and polyprotein translation. The RNA translation takes place in the ER and is initiated by binding the 5'UTR IRES to the ribosome, leading to the formation of the polyprotein precursor.
The polyprotein is co- and post-translationally cleaved by the host or viral proteases to yield the structural proteins (core, E1, and E2) and the nonstructural proteins (p7, NS2, NS3, NS4A, NS4B, NS5A, and NS5B proteins). HCV RNA replication occurs within the membranous web (MW) in the ER and is catalyzed by the NS5B RNA-dependent RNA polymerase. The newly generated viral RNA is transported to the site of core assembly and then is encapsulated to form the nucleocapsid.
The viral nucleocapsids subsequently interact with the viral E1/E2 proteins (envelopment) and bud into the ER lumen and then are translocated to the Golgi. The viral virions finish maturation in the Golgi and are transported to the plasma membrane through very-low-density lipoprotein (VLDL) pathway and released from host hepatocytes.
HDV is a satellite RNA virus that depends on HBV surface antigen (HBsAg) for viral entry, assembly, and transmission.
The HDV genome encodes hepatitis delta antigen (HDAg), which exists in two forms: small HDAg and large HDAg.
HDV research commonly focuses on HBV-dependent infection, HDAg function, viral RNA replication, and HBV-HDV co-infection mechanisms.
Rizzetto et al. initially discovered HDV in HBV-infected individuals with severe hepatitis in 1977 [13]. HDV is a hybrid virus as it uses Hepatitis B surface antigen (HBsAg) as its envelope protein. This characteristic allows HDV to infect only patients that concomitantly carry HBV.
HDV particle is composed of an outer lipoprotein envelope made of the surface antigen of the HBV (HBsAg) and an inner ribonucleoprotein structure in which the HDV genome resides. The HDV genome is about 1.7 kb and encodes only for the hepatitis delta antigen (HDAg) with two forms: a 27 kDa large-HDAg (delta-Ag-L), and a small-HDAg of 24 kDa (delta-Ag-S) [14].
| HDV Proteins | Function |
|---|---|
| Large delta antigen (L-HDAg) | Crucial for viral assembly |
| Small delta antigen (S-HDAg) | Necessary to initiate and maintain HDV RNA replication |
HDV infection is dependent on the presence of the HBV as it requires HBV's envelope proteins for its entry, assembly, and transmission. HDV virions initially attach to HSPGs on the hepatocyte surface and subsequently enter host cells through NTCP receptor [15]. After viral envelope fusion with the endosomal membrane, HDV ribonucleoprotein (RNP) is released from the endosome and then transported to the cellular nucleus.
In the nucleolus, HDV RNA undergoes replication through a double rolling circle amplification mechanism, generating both antigenomic RNA and additional genomic RNA. After export to the cytoplasm, the mRNA is translated at the ER to generate HDAg proteins. HDAg proteins then re-enter the nucleus, where the S-HDAg enhances genome replication. S-HDAg and L-HDAg associate with new synthesized genomic RNA to form new RNPs.
The resulting RNPs are released to the cytoplasm, where L-HDAg aids in linkage to HBsAg in the ER to assemble new viral particles. The newly synthesized viral particles are subsequently released from the hepatocytes through the Golgi to infect adjacent cells.
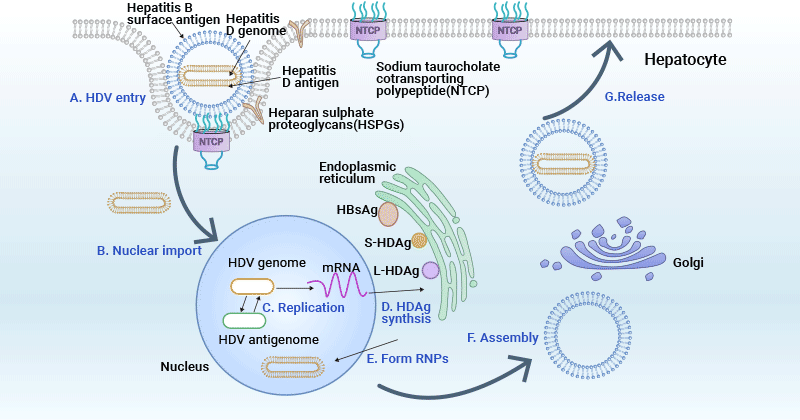
Figure 3. HDV life cycle
This picture is cited from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8557527/
HEV is a positive-sense single-stranded RNA virus mainly associated with fecal–oral, zoonotic, and blood transfusion-related transmission routes.
The HEV genome contains ORFs that encode non-structural proteins, capsid protein, and proteins involved in virion morphogenesis and release.
HEV-related research often focuses on viral entry, ORF protein function, replication, quasi-enveloped particles, and host-virus interaction.
Similar to HAV, HEV virions exist in two forms in the infected host, non-enveloped (neHEV) spherical particles in feces and quasi-enveloped (eHEV) particles in circulating blood and supernatant of infected cell cultures.
The HEV genomic RNA is about 7.2 kb in length and has three open reading frames (ORFs): ORF1 encodes a nonstructural polyprotein for virus replication and transcription, ORF2 encodes the capsid protein that contains the S, M, and P domains, and ORF3, which partially overlaps ORF2, encodes a multifunctional phosphoprotein involved in virion morphogenesis and pathogenesis.
Recently, a novel ORF4 has been discovered only in genotype 1 HEV, and endoplasmic reticulum (ER) stress boosts viral replication by promoting the translation of the novel ORF4 in genotype 1 HEV [16].
| HEV Proteins | Function |
|---|---|
| ORF1 | Encodes a non-structural polyprotein for virus replication and transcription |
| ORF2 | Encodes the capsid structural protein that is highly immunogenic and elicits neutralizing antibodies |
| ORF3 | Encodes a multifunctional phosphoprotein harboring two hydrophobic domains (D1 and D2) and two proline-rich domains (P1 and P2) |
| ORF4 | Only identified in genotype 1 HEV |
HEV infection begins when an individual consumes feces-contaminated water or food, introducing the virus into the digestive system. HEV subsequently enters the bloodstream through viremia and reaches the liver. Non-enveloped HEV particles bind to HSPGs on the surface of hepatocytes and enter host cells via a specific cellular receptor. Quasi-enveloped HEV virions enter liver cells through dynamin-dependent, clathrin-mediated endocytosis, involving Rab5 and Rab7.
After entry into host hepatocytes, HEV particles undergo uncoating of capsid protein and then release its genomic RNA to the cytoplasm. The viral genomic RNA serves directly as mRNA for the translation of ORF1 polyprotein. It also synthesizes a complementary negative-sense RNA, which acts as a template for the transcription of genomic and subgenomic mRNA.
The subgenomic mRNA is then translated to ORF2 capsid protein and ORF3 multifunctional phosphoprotein. ORF2 capsid protein forms virus-like particles (VLPs) and packages newly synthesized positive-sense genomic RNA to produce progeny HEV virions.
ORF3 modulates the host environment by interacting with cellular proteins, facilitating viral replication and release. Notably, ORF3 associates with TSG101 in the ESCRT pathway, aiding the budding of nascent virions into multivesicular bodies. These multivesicular bodies fuse with the plasma membrane, releasing viral virions from infected hepatocytes, either into the bloodstream as eHEV or in the bile duct as neHEV.
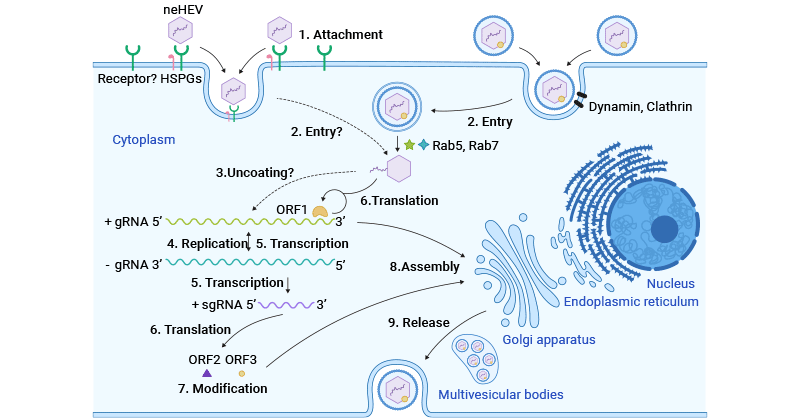
Figure 3. HEV life cycle
This picture is cited from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8079827/
The global situation of hepatitis viruses remains a significant public health concern. Hepatitis B and C, in particular, pose ongoing challenges due to their ability to cause chronic infections leading to severe liver diseases. The development of effective antiviral therapies and vaccination programs has made progress in preventing and managing hepatitis infections. However, barriers to access, especially in low-income regions, persist.
| Target Name | Product Name | Species Reactivity | Tested Applications | Code |
|---|---|---|---|---|
| P | P Antibody | Hepatitis B virus genotype A2 subtype adw2 | ELISA | CSB-PA324598LA01HDO |
| X | X Antibody | Hepatitis B virus genotype D | ELISA | CSB-PA526046LA01HVP |
| X | X Antibody | Hepatitis B virus genotype D subtype ayw | ELISA | CSB-PA865559LA01HEO |
| X | X Antibody | Hepatitis B virus genotype D subtype ayw | ELISA | CSB-PA355969LA01HEM |
| Genome polyprotein | Genome polyprotein Antibody | Hepatitis C virus genotype 1a | ELISA | CSB-PA333180LA01HFD |
| ORF1 | ORF1 Antibody | Hepatitis E virus genotype 1 | ELISA | CSB-PA327230LA01HGE |
| ORF2 | ORF2 Antibody | Hepatitis E virus genotype 1 | ELISA | CSB-PA727851LA01HVZ |
| ORF3 | ORF3 Antibody | Hepatitis E virus genotype 1 | ELISA | CSB-PA527126LA01HVZ |
Q:What are the major types of hepatitis viruses?
The major hepatitis viruses commonly discussed in research are hepatitis A virus (HAV), hepatitis B virus (HBV), hepatitis C virus (HCV), hepatitis D virus (HDV), and hepatitis E virus (HEV). These viruses differ in genome type, transmission route, viral protein composition, and infection outcome.
Q:Which hepatitis virus is a DNA virus?
Among HAV, HBV, HCV, HDV, and HEV, hepatitis B virus (HBV) is the only DNA virus. HBV contains a partially double-stranded relaxed circular DNA genome, while the other major hepatitis viruses are RNA viruses.
Q:How do HAV and HEV differ from HBV, HCV, and HDV?
HAV and HEV are commonly associated with fecal–oral transmission, often through contaminated food or water. HBV, HCV, and HDV are mainly associated with exposure to infected blood or body fluids. These differences influence how each virus is studied in virology and infectious disease research.
Q:Why is HDV dependent on HBV?
Hepatitis D virus (HDV) requires hepatitis B surface antigen (HBsAg) from HBV for viral entry, assembly, and transmission. Because of this dependence, HDV infection occurs in the context of HBV infection.
Q:What are key HBV proteins studied in research?
Common HBV proteins studied in research include hepatitis B surface antigen (HBsAg), hepatitis B core antigen (HBcAg), hepatitis B e antigen (HBeAg), HBx protein, and HBV DNA polymerase. These proteins are involved in viral structure, replication, immune response, and host-virus interaction studies.
Q:What are key HCV proteins studied in research?
HCV encodes a polyprotein that is processed into structural and non-structural proteins. Key HCV proteins studied in research include core protein, envelope glycoproteins E1 and E2, NS3, NS4A, NS4B, NS5A, and NS5B. These proteins are commonly investigated in viral entry, replication, assembly, and host interaction studies.
Q:What types of hepatitis virus research products does CUSABIO provide?
CUSABIO provides hepatitis virus-related research products, including recombinant viral proteins, antibodies, and ELISA kits. These products support studies involving viral protein function, host-virus interaction, immune response, and infectious disease mechanisms.
Q:Can recombinant hepatitis virus proteins be used for antibody development or assay development?
Recombinant hepatitis virus proteins can be used in research applications such as antigen-antibody interaction studies, antibody screening, assay development, and viral protein function research. Product suitability depends on the specific antigen, expression system, tag, purity, and intended experimental design.
Q:Are CUSABIO hepatitis virus-related products intended for clinical or diagnostic use?
No. CUSABIO hepatitis virus-related products listed on this page are intended for research use only. They are not intended for clinical use, diagnostic procedures, or therapeutic applications.
Q:How should researchers choose hepatitis virus-related products?
Researchers should choose products based on the virus type, target protein, genotype or subtype, application, sample type, species reactivity, expression system, and validation data. For studies involving viral proteins, it is also important to consider whether the target is structural, non-structural, or involved in viral entry, replication, assembly, or immune response.
References:
[1] Gholizadeh O, Akbarzadeh S, et al. Hepatitis A: Viral Structure, Classification, Life Cycle, Clinical Symptoms, Diagnosis Error, and Vaccination [J]. Can J Infect Dis Med Microbiol. 2023 Jan 4;2023:4263309.
[2] A. Schulze, P. Gripon, and S. Urban. Hepatitis B virus infection initiates with a large surface protein-dependent binding to heparan sulfate proteoglycans [J]. Hepatology, 46 (2007), pp. 1759-1768.
[3] E.R. Verrier, C.C. Colpitts, et al. A targeted functional RNA interference screen uncovers glypican 5 as an entry factor for hepatitis B and D viruses [J]. Hepatology, 63 (2016), pp. 35-48.
[4] H. Yan, G. Zhong, eet al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus [J]. Elife, 1 (2012), Article e00049.
[5] M. Iwamoto, W. Saso, et al. Epidermal growth factor receptor is a host-entry cofactor triggering hepatitis B virus internalization [J]. Proc. Natl. Acad. Sci. U. S. A., 116 (2019), pp. 8487-8492.
[6] Seeger C, Mason WS. Molecular Biology of Hepatitis B Virus Infection [J]. Virology (2015) 479-480:672–86.
[7] Schreiner S, Nassal M. A Role for the Host DNA Damage Response in Hepatitis B Virus cccDNA Formation-And Beyond [J]? Viruses (2017) 9(5):125.
[8] Tong S, Revill P. Overview of Hepatitis B Viral Replication and Genetic Variability [J]. J Hepatol (2016) 64(1 Suppl):S4–16.
[9] Molina S, Castet V, Pichard-Garcia L, et al. Serum-derived hepatitis C virus infection of primary human hepatocytes is tetraspanin CD81 dependent [J]. Journal of Virology. 2008;82:569-574.
[10] Scarselli E, Ansuini H, Cerino R, et al. The human scavenger receptor class B type I is a novel candidate receptor for the hepatitis C virus [J]. The EMBO Journal. 2002;21:5017-5025.
[11] Liu S, Yang W, Shen L, et al. Tight junction proteins claudin-1 and occludin control hepatitis C virus entry and are downregulated during infection to prevent superinfection [J]. Journal of Virology. 2009;83:2011-2014.
[12] Tscherne DM, Jones CT, et al. Time- and temperature-dependent activation of hepatitis C virus for low-pH-triggered entry [J]. J Virol. 2006;80:1734-1741.
[13] Rizzetto M, Canese MG, et al. Immunofluorescence detection of new antigen-antibody system (delta/anti-delta) associated to hepatitis B virus in liver and in serum of HBsAg carriers [J]. Gut. 1977;18(12):997–1003.
[14] Weiner AJ, Choo QL, et al. A single antigenomic open reading frame of the hepatitis delta virus encodes the epitope(s) of both hepatitis delta antigen polypeptides p24 delta and p27 delta [J]. J Virol. 1988;62(2):594–9.
[15] Li J, Wands J. Hepatitis B and D viral receptors [J]. Hepatology. 2016;63(1):11–3.
[16] V.P. Nair, S. Anang, C. Subramani, A. Madhvi, K. Bakshi, A. Srivastava, et al. Endoplasmic Reticulum Stress Induced Synthesis of a Novel Viral Factor Mediates Efficient Replication of Genotype-1 Hepatitis E Virus [J]. PLoS Pathog, 12 (2016), Article e1005521.
Proteins
Antibodies
ELISA Kits