Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-E04639m |
| Size | 96T,5×96T,10×96T |
| Price | Request a Quote |
| Trial Size |
24T ELISA Kit Trial Size (Only USD$150/ kit) * Sample kit cost can be deducted as a $30 credit for each 96-assay kit of the same analyte and brand you subsequently purchase within six months until depleted. More details >> Interested in a trial size? Please leave a message below.
|
| Have Questions? | Leave a Message or Start an on-line Chat |
Interleukin-6 (IL-6) is a multifunctional cytokine that plays important roles in immune responses, inflammation, and hematopoiesis. Originally identified as a B-cell hybridoma growth factor, IL-6 regulates the acute phase response, promotes B-cell differentiation into antibody-producing plasma cells, and stimulates T-cell activation. Various cell types produce this cytokine, including macrophages, T-cells, endothelial cells, and fibroblasts in response to inflammatory stimuli. IL-6 signaling occurs through binding to its receptor complex, triggering downstream pathways that influence cell proliferation, differentiation, and survival. This makes it an important mediator in both physiological and pathological processes.
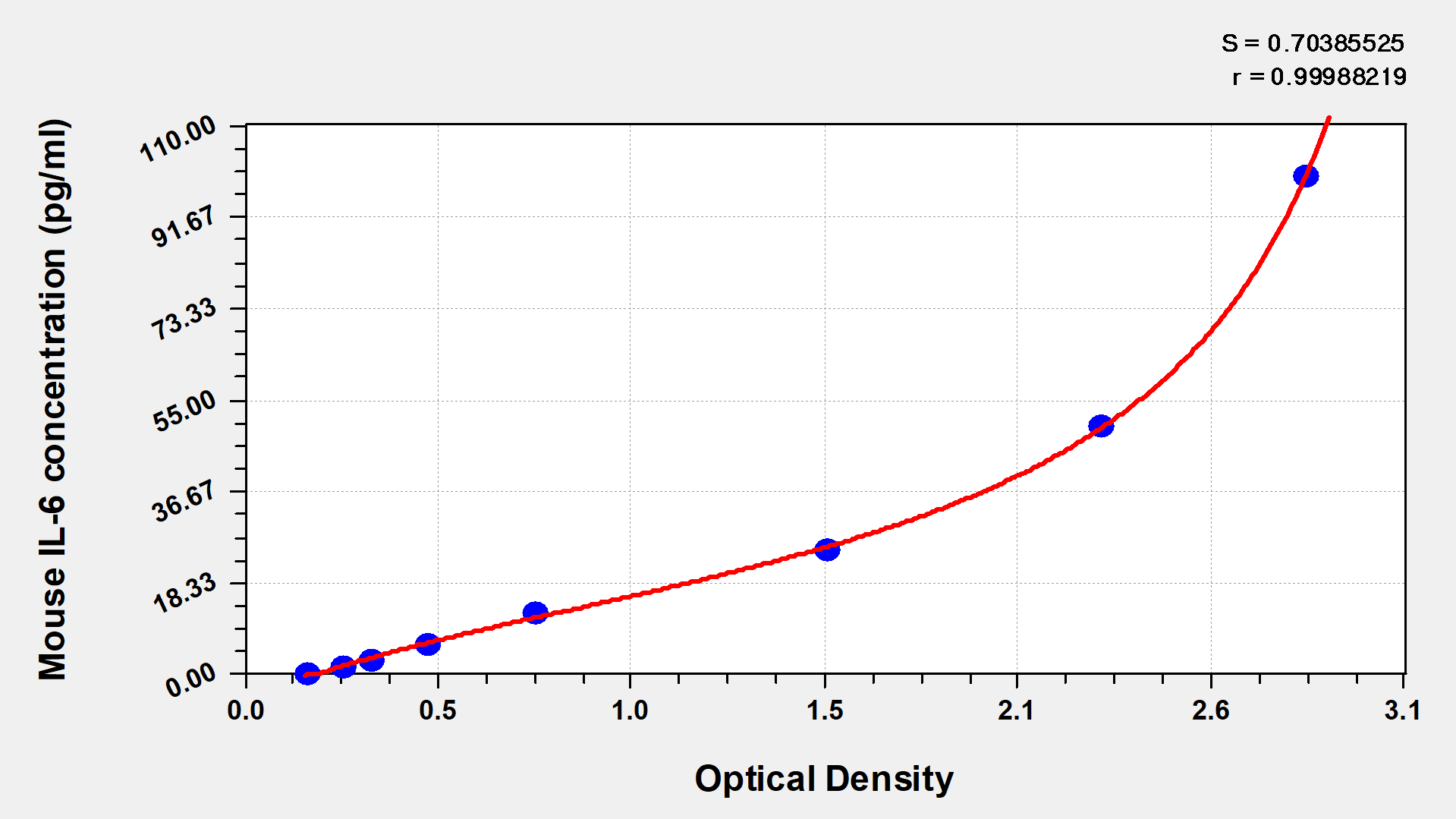
The Mouse Interleukin 6,IL-6 ELISA KIT (CSB-E04639m) is designed for quantitative detection of IL-6 in Mus musculus samples using a sandwich measurement approach. This assay requires 50-100ul sample volume and can be completed within 1-5 hours. Detection is performed at 450 nm wavelength, providing researchers with a reliable method for measuring mouse IL-6 levels in their experimental studies.
Application Examples
Note: The following application examples are drawn from a selection of publications citing this product. For additional applications, please refer to the full list of references in the "Citations" section.
This mouse IL-6 ELISA kit has been used in inflammatory research to quantify IL-6 levels in various biological samples. The kit supports studies that examine cytokine responses in cellular and animal models, particularly in the context of immune activation and inflammatory processes.
• Inflammatory cytokine profiling - Measurement of IL-6 alongside other pro-inflammatory cytokines including TNF-α and IL-1β in comprehensive cytokine panels
• Macrophage research - Quantification of IL-6 secretion from cultured bone marrow-derived macrophages following various treatments and stimulations
• Plasma biomarker analysis - Measurement of circulating IL-6 levels in mouse plasma samples collected from experimental animal models
• Cell culture supernatant analysis - Evaluation of IL-6 production in cell culture systems to study inflammatory responses and cytokine release
• Gastrointestinal inflammation studies - Examination of inflammatory markers in experimental colitis models using dextran sodium sulfate
• Pharmacological intervention research - Measurement of IL-6 modulation in response to various therapeutic compounds and experimental treatments
• NLRP3 inflammasome research - Study of IL-6 production in the context of inflammasome activation and related inflammatory pathways
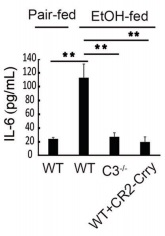
Sample type: Serum
Sample species: Mouse
Review: In addition, TNF-α and IL-6 levels were increased in the ethanol-fed mice, and CR2-Crry treatment significantly reduced the levels of both cytokines.
By Anonymous
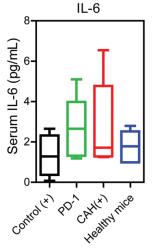
Sample type: Serum
Sample species: Mouse
Review: Serum proinflammatory cytokines levels including interleukin 2 (IL-2), interleukin 6 (IL-6), tumor necrosis factor α (TNF-α) and interferon gamma (IFN-γ) were measured after two-cycle treatments.
By Anonymous
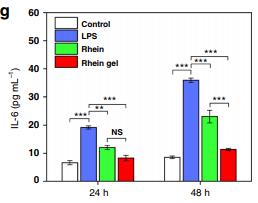
Sample type: Cell culture supernatant
Sample species: Mouse
Review: BV2 cells (1 × 105 cells per well in a 24-well plate) were pretreated with treatments for 1 h and stimulated with LPS (100 ng mL−1). After treatment for 24 h and 48 h, the supernatants were collected. The concentrations of TNF-α, IL-1β, IL-6, IL-12, and iNOS in the culture medium were measured by ELISA kits.
By Anonymous
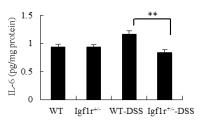
Sample type: Cell culture supernatant
Sample species: Mouse
Review: Levels of TNF-α (CSB-E04741m), IL-6 (CSB-E04639m) and PGE2 (CSB-E07966m) in supernatant of colorectal mucosa isolated from mice were quantified by using ELISA kits according to the manufacturer\'s protocols. ELISA analysis of IL-6, TNF-α and PGE2 in the intraepithelial and lamina propria showed significant differences between WT and Igf1r+/- mice.
By Anonymous
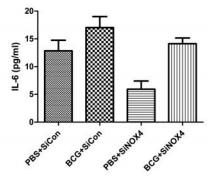
Sample type: Biological fluids(Pleural fluid)
Sample species: Mouse
Review: The presence of interferon-γ, IL-2, TNF-α, monocyte chemotactic protein (MCP) -1, and IL-6 in pleural tissue was quantified using an enzyme-linked immunosorbent assay. The concentration of cytokines (IL-6, TNFA, IFN, MCP-1, IL-2) in pleural effusion was higher in mice in the BCG+ SiCon group than in the PBS+ SiCon group.
By Anonymous
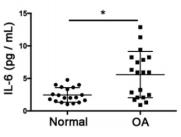
Sample type: Plasma (anticoagulant)
Sample species: Mouse
Review: The results of ELISA assays indicated that the expression levels of common proinflammatory indicators, such as IL-6 and IL-8, were significantly higher in the OA group compared to those in the normal group.
By Anonymous
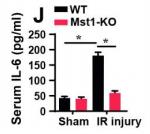
Sample type: Plasma (anticoagulant)
Sample species: Mouse
Review: ELISA was used to analyze the levels of inflammation factors after renal I/R injury.
By Anonymous
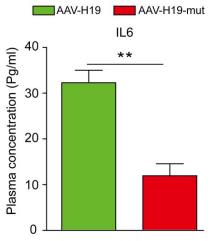
Sample type: Plasma (anticoagulant)
Sample species: Mouse
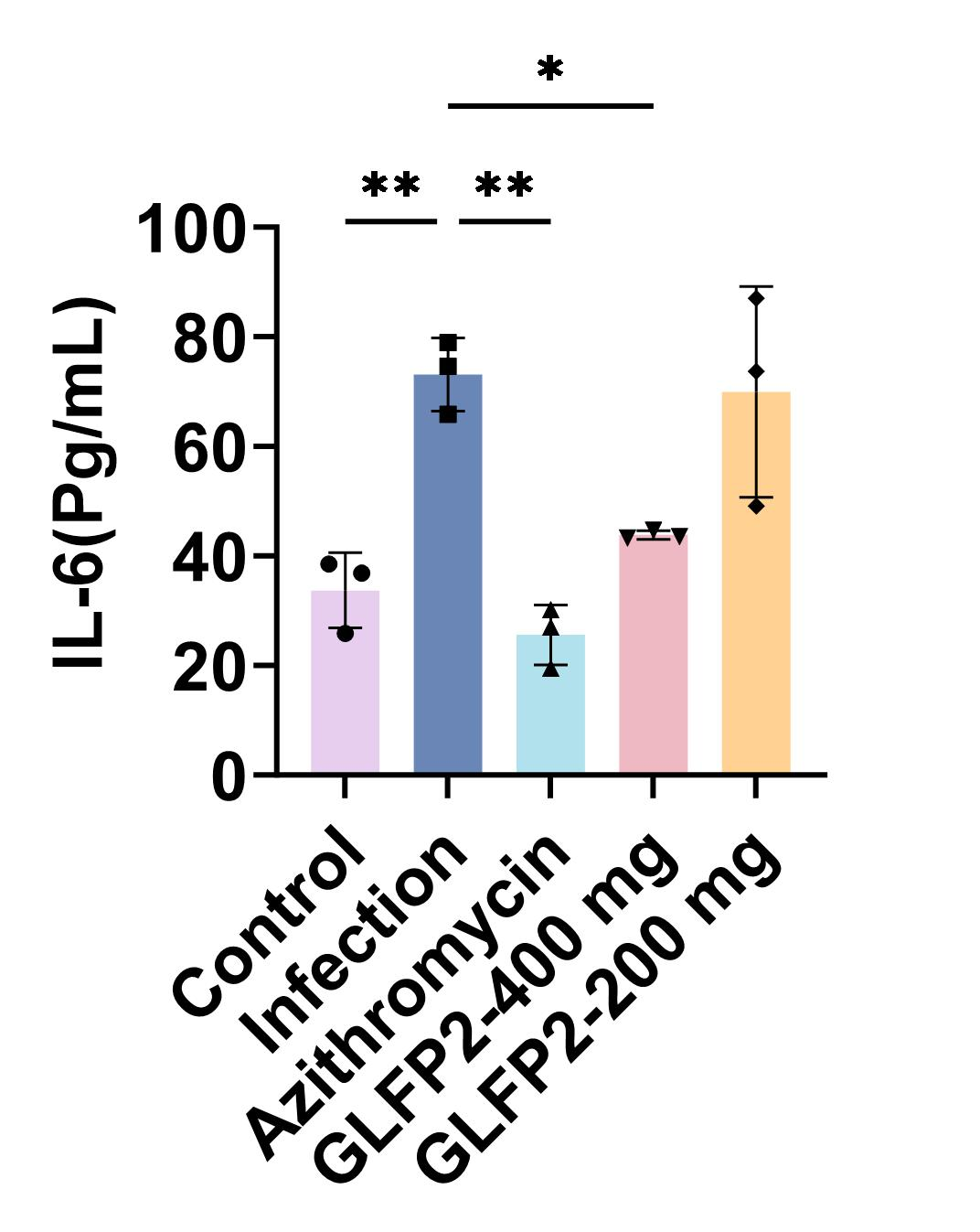
Review: IL-6 concentrations in mouse plasma as well as in ex vivo culture solution were measured using a mouse IL-6 ELISA kit (CSB-E04639m) according to the manufacturer’s instructions. Plasma IL-6 concentration (n=8 per group). Data are presented as the mean±SD or as median and IQR (for elastin degradation score). *P<0.05, **P<0.01.
By Anonymous
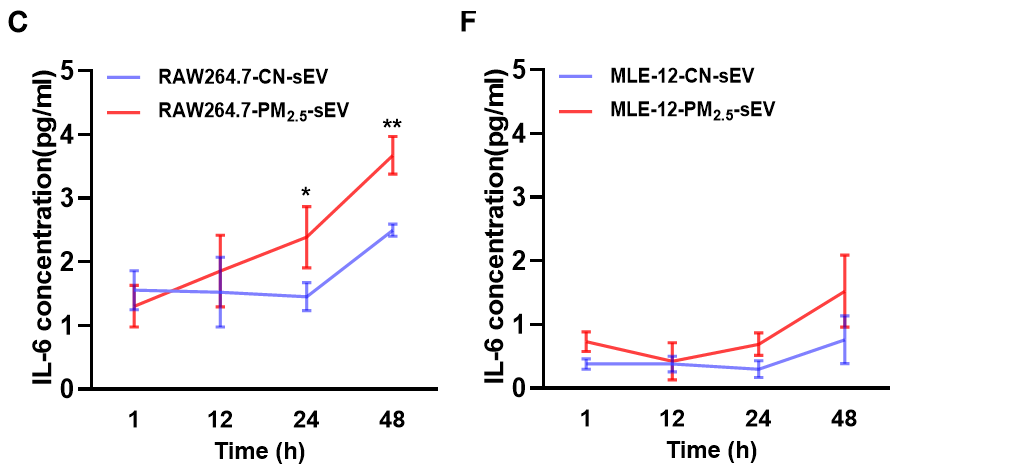
Sample type: Cell culture supernatant
Sample species: Mouse
Sample dilution: No dilution
Review: I used CSB-E04639m to detect IL6 levels in cell supernatant exosomes and mouse supernatants, respectively. The results of the standard curve were good, as shown in the Appendix. However, during the actual assay, probably due to the small changes in IL-6 in the treated group, the majority of values were below the detection limit, but some values could still be detected after increasing the sample size, and the final results could be used in the article.
By Anonymous
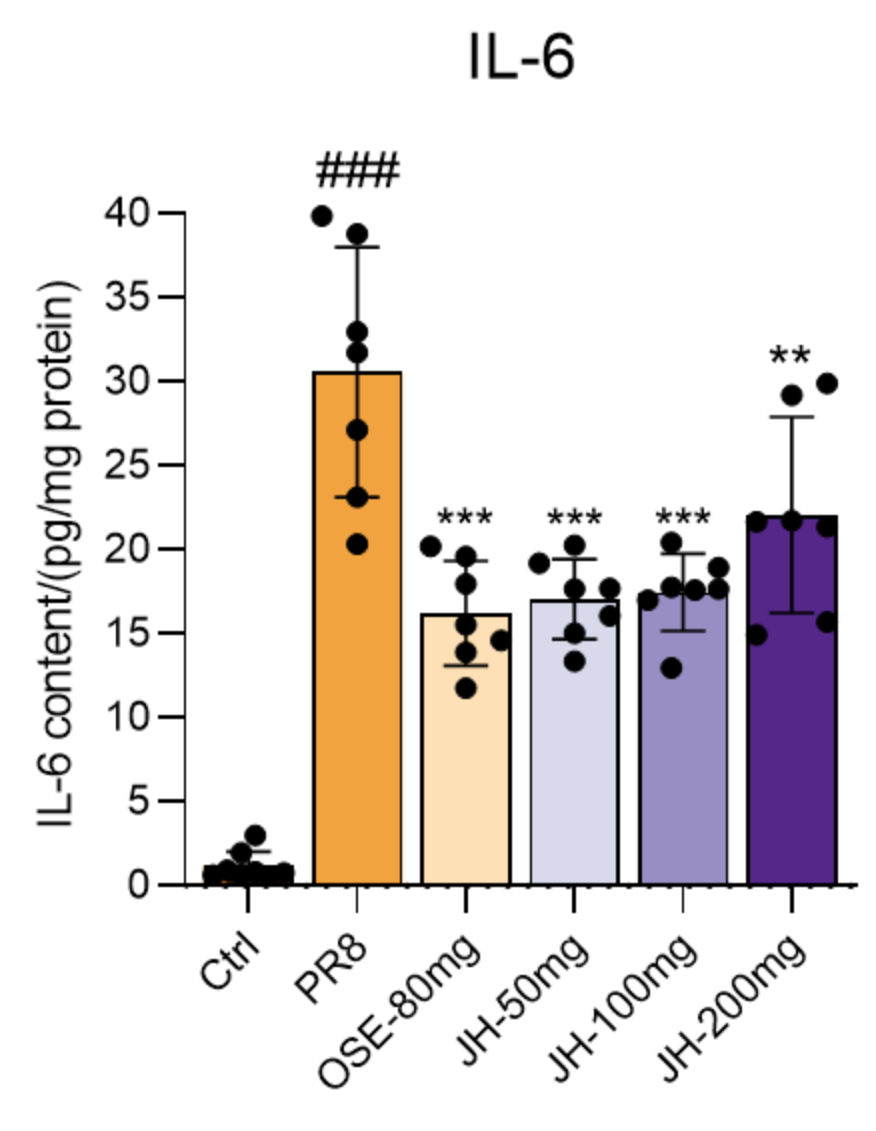
Sample type: Tissue homogenate
Sample species: Mouse
Sample dilution: No dilution
Review: I have tested the CSB-E04639m for PR8 virus-infected mouse lung tissue homogenates, the operation is simple and easy to use, the sensitivity is very high, and the test results are very accurate.
By Anonymous
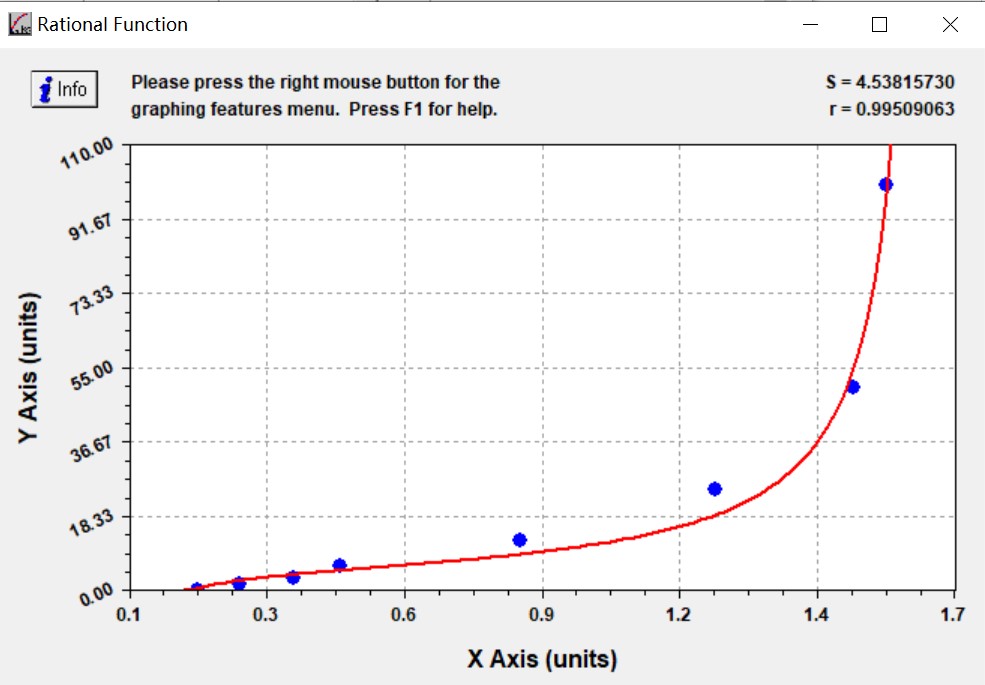
Sample type: Serum
Sample species: Mouse
Sample dilution: 1:100
Review: I used CSB-E04639m to detect spontaneous T2DM mice db/db mice, the dilution ratio was 1: 5, the dilution ratio was large, the concentration was low, the undiluted serum concentration was just right, and the standard curve of this product was particularly accurate!
By Anonymous
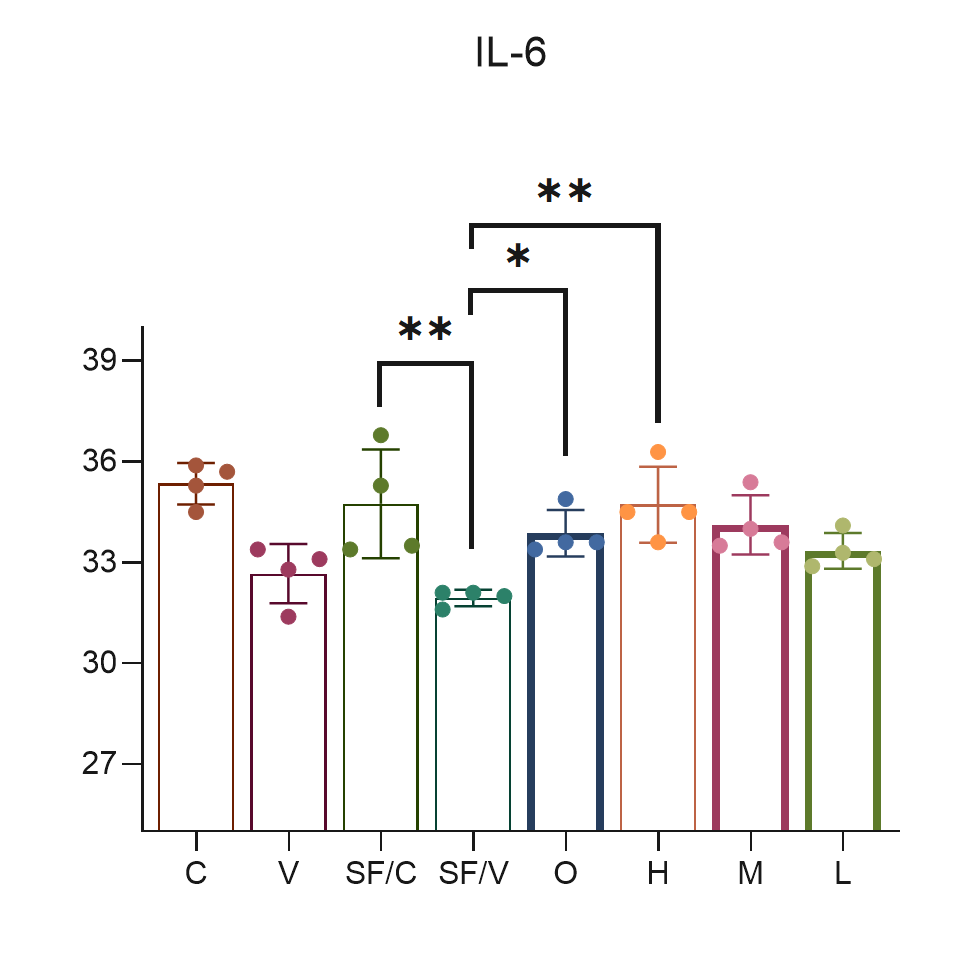
Sample type: Serum
Sample species: Mouse
Sample dilution: No dilution
Review: I have detected influenza virus infection in the peripheral blood supernatant of mice using CSB-E04639m. The operation is simple, convenient, and easy to get started, with very high sensitivity and very accurate detection results.
By Anonymous
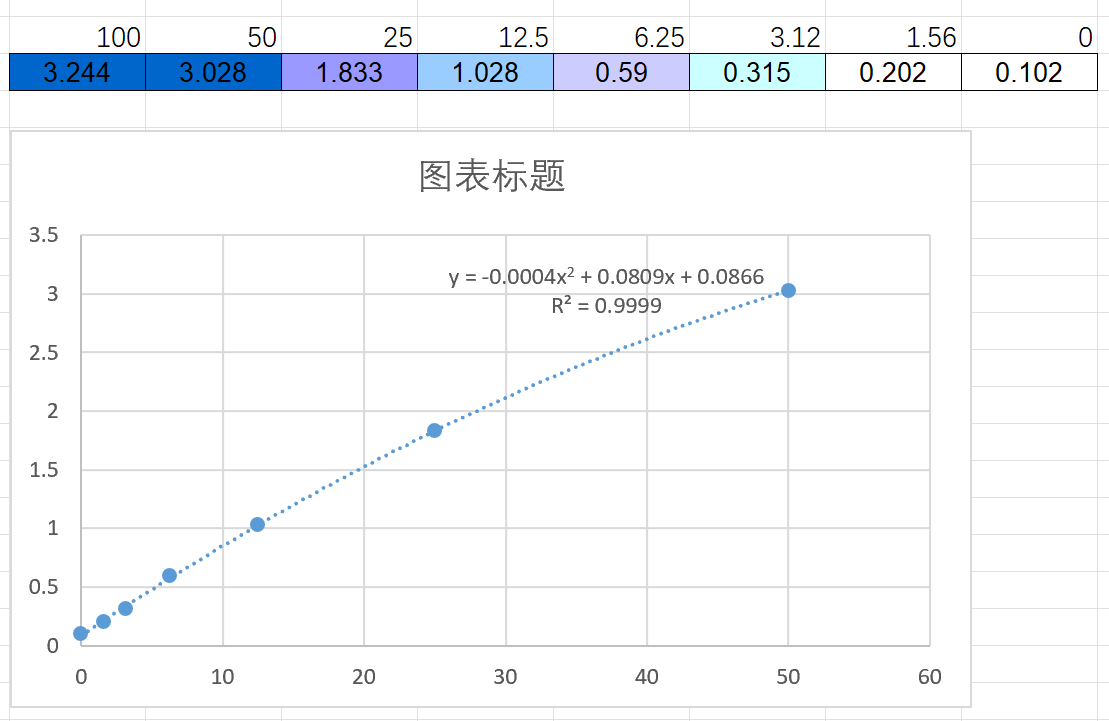
Sample type: Biological fluids(bronchoalveolar lavage fluid)
Sample species: Mouse
Sample dilution: No dilution
Review: We used CSB-E04639m to measure the IL-6 level in bronchoalveolar lavage fluid of normal and smoking C57B/L mice. The OD value of normal group was 0.049-0.478, and that of smoking group was 0.844-1.252.
By Anonymous
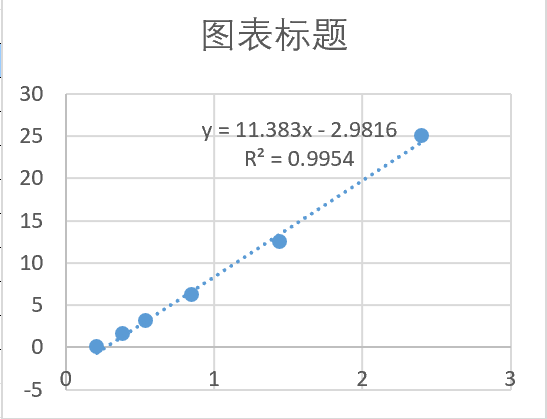
Sample type: Cell culture supernatant
Sample species: Mouse
Sample dilution: 1:2
Review: CSB-E04639m was used to test the culture supernatant of mouse DC2.4 cells. The standard curve showed that the od value of the control group was 0.46-0.49, and the OD value of the mature group stimulated with IFN was 0.43-0.5.
By Anonymous
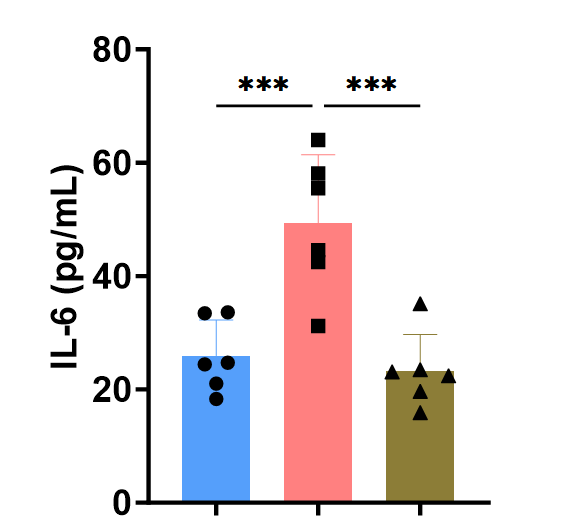
Sample type: Tissue homogenate Lung
Sample species: Mouse
Sample dilution: 1:50
Review: I used CSB-E04639m to detect the IL-6 level in mouse lung tissue homogenate before and after modeling and treatment. The standard curve result was good and consistent with the expected result. The kit is easy to operate, with simple and convenient steps, saving experimental time.
By Anonymous
Sample type: Tissue homogenate Lung
Sample species: Mouse
Sample dilution: 1:50
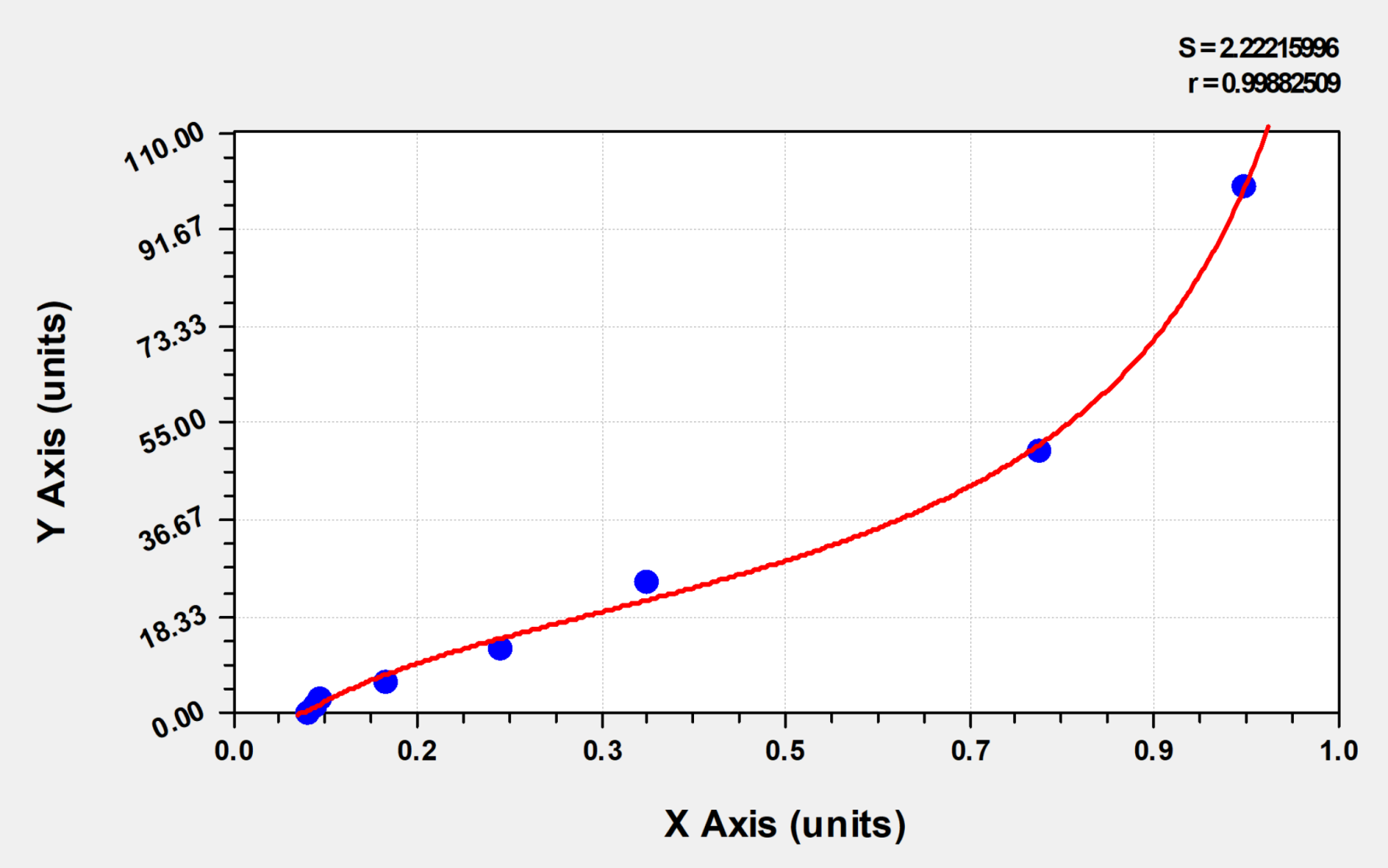
Review: Mycoplasma x mouse lung homogenate plasma was detected using CSB-E04639m with a 1:50 dilution. The sample OD value was 0.052, which was between the OD values of S2 and S3. The experiment was feasible, with little difference between replicate wells and within the group.
By Anonymous
Sample type: Serum
Sample species: Mouse
Sample dilution: 1:50
Review: I use the CSB-E04639m. The OD value range is 0.114-0.964. The product is sensitive, very easy to use, and takes less time. The results are very reliable. I often buy Huamei's ELISA products and hope they continue to maintain the quality.
By Anonymous

Sample type: Serum
Sample species: Mouse
Sample dilution: No dilution
Review: We used CSB-E04639m to detect the serum of normal and atherosclerotic mice. The OD value of the standard curve was 0-50, and the OD value of the samples was 2-10. Overall, the quality of the standard solution was stable, and the standard curve results were reliable. The sample detection results could also clearly distinguish different treatment groups, which was consistent with physiological and pathological cognition, indicating that the detection kit was credible. According to the volume of the sample we added (50 microliters), the amount of the standard we added was also halved. Therefore, the highest point of the standard curve was 50, and the response of the sample was 2-10, which was the actual detection result.
By Anonymous

