Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-EP011664HU |
| Abbreviation | Recombinant Human IL6 protein (Active) |
| MSDS | |
| Size | $224 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
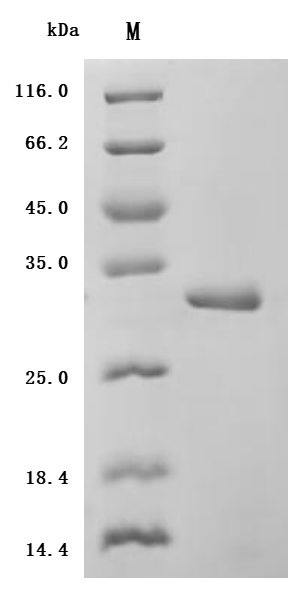
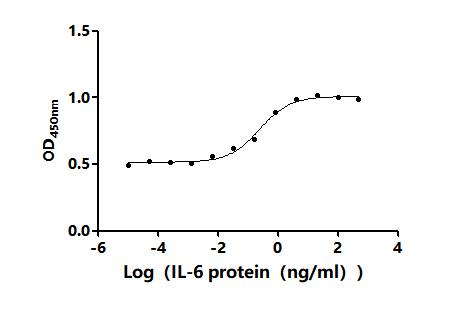
IL-6 is a pleiotropic cytokine central to JAK/STAT3 signaling, acute-phase responses, and tumor microenvironment biology, making precise functional activity essential for meaningful experimental results. This recombinant human IL-6, spanning the full mature sequence (residues 30–212) with an N-terminal 6×His tag, demonstrates potent bioactivity with an ED50 of 0.15–0.40 ng/mL in TF-1 proliferation assays, confirming signaling competence suitable for cell-based functional studies including proliferation, differentiation, and JAK/STAT or NF-κB pathway activation experiments. Purity exceeding 95% by SDS-PAGE and endotoxin levels below 1.0 EU/μg minimize LPS-driven artifacts that can confound immune cell assays, providing a suitable basis for in vivo tumor biology models and antibody development workflows such as ELISA standard curves or Western blot positive controls. The E. coli-derived, tag-accessible format also supports use as a reference standard in cytokine detection assays where batch-to-batch consistency matters.
There are currently no reviews for this product.

