Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)

On May 17, 2026, the WHO declared that the Ebola virus disease outbreaks in the Democratic Republic of the Congo and Uganda (Bundibugyo region) constituted a "Public Health Emergency of International Concern" (PHEIC). According to media reports, as of May 30 (local time), the DRC government updated the latest response status of the ongoing outbreak, reporting more than 260 confirmed cumulative cases. The epidemic has spread across multiple areas in Ituri Province, North Kivu Province, and South Kivu Province, with suspected cases also reported in several countries outside Africa.
Ebola virus (EBOV) is a member of the genus Ebolavirus in the family Filoviridae. It is the causative agent of Ebola virus disease (Ebola virus disease), also known as Ebola hemorrhagic fever.
Six species of Ebola virus are currently recognized:
Among these, Zaire ebolavirus, Sudan ebolavirus, and Bundibugyo ebolavirus have been responsible for major outbreaks. Taï Forest ebolavirus can cause human infection but has not led to sustained transmission. Reston and Bombali ebolaviruses are not associated with severe human disease.
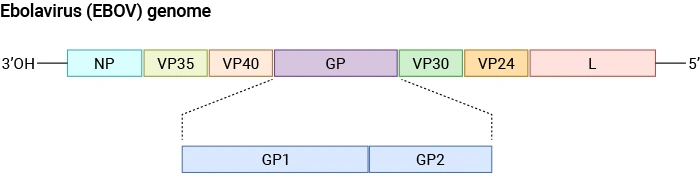
Schematic diagram of the Ebolavirus (EBOV) genome structure.
Source:PMID: 40733577
The pathogenicity of Ebola virus is driven by its highly efficient cell entry, potent suppression of host immunity, and dysregulated inflammatory responses leading to systemic tissue damage and multi-organ failure.
The virus enters the body through breaches in the skin, infecting epidermal keratinocytes and exploiting the AXL receptor to facilitate entry. It then rapidly targets macrophages and dendritic cells, spreading to lymph nodes, liver, and spleen, ultimately resulting in systemic dissemination [1].
At the immune level, the viral protein VP35 suppresses type I interferon responses through multiple mechanisms, while also hijacking the PKA–CREB1 pathway to enhance replication and upregulate coagulation-related gene expression. Despite interferon suppression, infection triggers a strong inflammatory response: VP40 activates the NF-κB pathway, inducing massive pro-inflammatory cytokine release and a cytokine storm. It also promotes apoptosis of bystander lymphocytes via exosome-mediated signaling, further exacerbating immunosuppression [2].
Vascular pathology includes endothelial dysfunction, capillary leakage, and coagulation abnormalities, progressing to hypovolemic shock and disseminated intravascular coagulation (DIC). The virus disseminates widely to organs such as the liver, lungs, and kidneys, leading to multi-organ failure. Host genetic factors also contribute: TRIM5 loci are associated with liver failure risk, whereas TRIM14 has antiviral protective effects [3].
In addition, EBOV can persist in immune-privileged sites (e.g., aqueous humor of the eye), leading to delayed complications. Viral miRNA-like molecules and secreted δ-peptides may further modulate host signaling pathways and immune responses, highlighting a complex virus–host interaction network [4].
A hallmark of fatal Ebola virus infection is profound immune dysregulation. Compared with survivors, deceased patients often exhibit more intense upregulation of interferon and inflammatory pathways during the acute phase.
Overall, Ebola virus pathogenesis is a dynamic, multi-layered process ranging from molecular hijacking (e.g., VP35 and VP40 functions), to cellular destruction, and ultimately systemic immune dysregulation, coagulopathy, and multi-organ failure. Understanding host susceptibility loci (e.g., TRIM5) and protective factors (e.g., TRIM14), as well as viral persistence mechanisms, provides critical targets for therapeutic and vaccine development.
CUSABIO offers the following products for Ebola virus research.
| Target | Product Name | Code | Tag Info | Source |
|---|---|---|---|---|
| GP | Recombinant Zaire ebolavirus Super small secreted glycoprotein (GP) | CSB-BP314099ZAT | N-terminal 10xHis-tagged and C-terminal Myc-tagged | Baculovirus |
| VP24 | Recombinant Zaire ebolavirus Membrane-associated protein VP24 (VP24) | CSB-BP522532ZAS | N-terminal 10xHis-tagged and C-terminal Myc-tagged | Baculovirus |
| VP40 | Recombinant Zaire ebolavirus Matrix protein VP40 (VP40) | CSB-BP762349ZAT | N-terminal 10xHis-tagged and C-terminal Myc-tagged | Baculovirus |
| GP | Recombinant Zaire ebolavirus Pre-small/secreted glycoprotein (GP) | CSB-EP308957ZAA | N-terminal 6xHis-tagged | E.coli |
| GP | Recombinant Zaire ebolavirus Envelope glycoprotein (GP), partial | CSB-EP311249ZAT | N-terminal 6xHis-tagged | E.coli |
| L | Recombinant Zaire ebolavirus RNA-directed RNA polymerase L (L), partial | CSB-EP313164ZAB | N-terminal 6xHis-tagged | E.coli |
| L | Recombinant Zaire ebolavirus RNA-directed RNA polymerase L (L), partial | CSB-EP313164ZABb1 | N-terminal 10xHis-tagged and C-terminal Myc-tagged | E.coli |
| GP | Recombinant Zaire ebolavirus Super small secreted glycoprotein (GP) | CSB-EP314099ZAT | N-terminal 6xHis-SUMO-tagged | E.coli |
| NP | Recombinant Zaire ebolavirus Nucleoprotein (NP), partial | CSB-EP323576ZAB | N-terminal 6xHis-tagged | E.coli |
| NP | Recombinant Zaire ebolavirus Nucleoprotein (NP), partial | CSB-EP323576ZABa2 | N-terminal 6xHis-SUMO-tagged | E.coli |
| GP | Recombinant Zaire ebolavirus Pre-small/secreted glycoprotein (GP) | CSB-EP350772ZAT | N-terminal 6xHis-SUMO-tagged | E.coli |
| VP24 | Recombinant Zaire ebolavirus Membrane-associated protein VP24 (VP24) | CSB-EP522532ZAS | N-terminal 10xHis-tagged and C-terminal Myc-tagged | E.coli |
| VP30 | Recombinant Zaire ebolavirus Minor nucleoprotein VP30 (VP30) | CSB-EP755264ZAT | N-terminal 6xHis-SUMO-tagged | E.coli |
| VP30 | Recombinant Zaire ebolavirus Minor nucleoprotein VP30 (VP30) | CSB-EP755264ZATa0 | N-terminal 6xHis-tagged | E.coli |
| VP40 | Recombinant Zaire ebolavirus Matrix protein VP40 (VP40) | CSB-EP762349ZAT | N-terminal 6xHis-SUMO-tagged | E.coli |
| VP40 | Recombinant Zaire ebolavirus Matrix protein VP40 (VP40) | CSB-EP762349ZATa0 | N-terminal 6xHis-tagged | E.coli |
| L | Recombinant Zaire ebolavirus RNA-directed RNA polymerase L (L), partial | CSB-EP764949ZAT | N-terminal 10xHis-tagged and C-terminal Myc-tagged | E.coli |
| VP35 | Recombinant Zaire ebolavirus Polymerase cofactor VP35 (VP35) | CSB-EP764950ZAT | N-terminal 10xHis-tagged and C-terminal Myc-tagged | E.coli |
| VP40 | Recombinant Reston ebolavirus Matrix protein VP40 (VP40) | CSB-EP810348RCJ | N-terminal 6xHis-SUMO-tagged | E.coli |
| NP | Recombinant Reston ebolavirus Nucleoprotein (NP), partial | CSB-EP841983RCI | N-terminal 6xHis-tagged | E.coli |
| NP | Recombinant Reston ebolavirus Nucleoprotein (NP), partial | CSB-EP841983RCIa2 | N-terminal 6xHis-SUMO-tagged | E.coli |
| NP | Recombinant Sudan ebolavirus Nucleoprotein (NP), partial | CSB-EP882531SRC | N-terminal 6xHis-tagged | E.coli |
| GP | Recombinant Zaire ebolavirus Super small secreted glycoprotein (GP) | CSB-MP314099ZAT | N-terminal 10xHis-tagged and C-terminal Myc-tagged | Mammalian cell |
| GP | Recombinant Sudan ebolavirus Envelope glycoprotein (GP), partial | CSB-MP742487SRE | N-terminal 6xHis-Myc-tagged | Mammalian cell |
| VP35 | Recombinant Zaire ebolavirus Polymerase cofactor VP35 (VP35) | CSB-MP764950ZAT | N-terminal 10xHis-tagged and C-terminal Myc-tagged | Mammalian cell |
| GP | Recombinant Zaire ebolavirus Pre-small/secreted glycoprotein (GP) | CSB-YP308957ZAA | N-terminal 6xHis-tagged | Yeast |
| GP | Recombinant Zaire ebolavirus Envelope glycoprotein (GP), partial | CSB-YP310843ZAA | N-terminal 6xHis-tagged | Yeast |
| GP | Recombinant Zaire ebolavirus Super small secreted glycoprotein (GP) | CSB-YP314099ZAT | N-terminal 6xHis-tagged and C-terminal Myc-tagged | Yeast |
| NP | Recombinant Zaire ebolavirus Nucleoprotein (NP), partial | CSB-YP323576ZAB | N-terminal 6xHis-tagged | Yeast |
| GP | Recombinant Zaire ebolavirus Pre-small/secreted glycoprotein (GP) | CSB-YP350772ZAT | N-terminal 6xHis-tagged | Yeast |
| GP | Recombinant Sudan ebolavirus Envelope glycoprotein (GP), partial | CSB-YP742487SRE | N-terminal 6xHis-tagged | Yeast |
| VP30 | Recombinant Zaire ebolavirus Minor nucleoprotein VP30 (VP30) | CSB-YP755264ZAT | N-terminal 6xHis-tagged | Yeast |
| GP | Recombinant Sudan ebolavirus Envelope glycoprotein (GP), partial | CSB-EP742487SRE | N-terminal 6xHis-KSI-tagged | E.coli |
| GP | Recombinant Reston ebolavirus Pre-small/secreted glycoprotein (GP), partial | CSB-EP852740RCI | N-terminal 6xHis-tagged | E.coli |
| GP | Recombinant Reston ebolavirus Pre-small/secreted glycoprotein (GP), partial | CSB-MP852740RCI | N-terminal 6xHis-tagged | Mammalian cell |
| Target | Product Name | Code | Antigen Species | Tested Applications |
|---|---|---|---|---|
| GP | GP Antibody | CSB-PA310843LA01ZAA | Zaire ebolavirus (strain Eckron-76) (ZEBOV) (Zaire Ebola virus) | ELISA |
| GP | GP Antibody | CSB-PA314099LA01ZAT | Zaire ebolavirus (strain Kikwit-95) (ZEBOV) (Zaire Ebola virus) | ELISA |
| GP | GP Antibody | CSB-PA350772LA01ZAT | Zaire ebolavirus | ELISA |
| VP40 | VP40 Antibody | CSB-PA810348HA01RCJ | Reston ebolavirus (strain Reston-89) (REBOV) (Reston Ebola virus) | ELISA |
| VP35 | VP35 Antibody | CSB-PA856262LA01RCT | Reston ebolavirus (strain Philippines-96) (REBOV) (Reston Ebola virus) | ELISA |
| GP | GP Antibody | CSB-PA308957LA01ZAA | Zaire ebolavirus | ELISA, WB |
| NP | NP Antibody | CSB-PA841983HA01RCI | Reston ebolavirus | ELISA, WB |
| VP40 | VP40 Antibody | CSB-PA762349HA01ZAT | Zaire ebolavirus | ELISA, WB |
Ebola virus is primarily transmitted through:
Blood and bodily fluids of infected individuals (vomit, feces, sweat, saliva, breast milk, semen).
Contaminated clothing, bedding, and medical equipment.
Fruit bats are considered natural reservoirs; humans can be infected through contact with infected wild animals (e.g., monkeys, gorillas).
Some studies suggest potential aerosol transmission under experimental conditions, but contact transmission remains the dominant route.
The incubation period ranges from 2 to 21 days, with an average of 8–10 days. Symptoms typically progress in stages:
High fever, fatigue, myalgia, headache, sore throat, nausea, vomiting
Diarrhea, abdominal pain, rash, persistent fever, bleeding tendency
Multi-organ failure, severe hemorrhage, hepatic and renal dysfunction; case fatality rates for certain strains may reach up to 90%
Currently, approved vaccines exist only for Zaire ebolavirus (e.g., Ervebo and Zabdeno/Mvabea). No licensed vaccines are available for Bundibugyo or Sudan ebolaviruses, although several candidates are under development.
Q:What are the genomic characteristics of EBOV, and why do pathogenicity differences exist among strains?
EBOV is a single-stranded negative-sense RNA virus with a genome of approximately 19 kb encoding seven structural proteins. The genome organization is: NP, VP35, VP40, GP, VP30, VP24, and L.
Differences in pathogenicity among species (e.g., Zaire vs. Bundibugyo) are primarily linked to variations in the glycoprotein (GP) and immune-modulating proteins VP35 and VP40. GP mediates receptor binding and membrane fusion, and its sequence variability directly affects viral entry efficiency. VP35 and VP40 function as key immune antagonists, suppressing type I interferon responses and promoting inflammatory signaling, respectively. Zaire ebolavirus has a case fatality rate of up to 90%, whereas Reston virus does not cause severe disease in humans.
Q:What are the key receptors and mechanisms of Ebola virus entry?
EBOV enters through skin breaches, initially infecting keratinocytes, and uses receptors such as AXL and TIM-1 to facilitate entry. Viral GP binds host receptors and enters cells via macropinocytosis.
Keratinocytes serve as early "landing cells," establishing local infection (Richards PT et al., J Virol, 2025). The virus then exploits macrophages and dendritic cells to disseminate systemically through the lymphatic network, enabling widespread infection.
Q:How does EBOV evade innate immunity and trigger a cytokine storm?
EBOV suppresses type I interferon responses via VP35, while VP40 activates NF-κB signaling, inducing excessive cytokine release. VP35 blocks RIG-I–like receptor pathways and inhibits IFN-β production, enabling early replication. VP40 drives production of TNF-α, IL-6, and other cytokines, and induces bystander lymphocyte apoptosis via exosomal signaling (Yamaoka S et al., PNAS, 2026).
This combination of interferon suppression and hyperinflammation is a central mechanism underlying the high lethality of Ebola virus disease. Anti-cytokine strategies (e.g., TNF-α neutralization) have been proposed as potential therapeutic approaches.
Q:How do host genetic factors influence disease progression?
Host genetics play a significant role in disease outcomes. TRIM5 polymorphisms are associated with liver failure risk, while TRIM14 exerts antiviral protective effects. TRIM proteins are E3 ubiquitin ligases involved in innate immune regulation.
TRIM5 variants may alter recognition of viral components and correlate with disease severity, whereas TRIM14 enhances interferon signaling (Falasca L et al., Cell Death Differ, 2015). These findings highlight the importance of genetic background in infection outcomes and suggest potential biomarkers for risk stratification and host-directed therapies (HDT).
Q:Does Ebola virus cause persistent infection? What are the implications for survivors?
EBOV can persist in immune-privileged sites such as the aqueous humor and seminal fluid, leading to delayed complications. Viral miRNA-like molecules and secreted δ-peptides may also modulate host pathways.
In some survivors, the virus can persist for months or longer, causing complications such as uveitis and arthralgia, and posing a risk of sexual transmission. These mechanisms illustrate immune evasion strategies that allow long-term persistence and have important implications for vaccination strategies and survivor management (Doghish AS et al., Folia Microbiol, 2026).
Q:What is the current status of Ebola vaccines and key antigen targets?
Currently approved vaccines exist only for Zaire ebolavirus (Ervebo; Zabdeno/Mvabea). No licensed vaccines are available for Bundibugyo or Sudan species. Glycoprotein (GP) and VP40 are the primary antigen targets in vaccine and antibody development.
Existing vaccines focus on GP to induce neutralizing antibodies. However, substantial sequence divergence across species limits cross-protection. VP40, involved in viral assembly and inflammatory signaling, is also a promising target. Current research focuses on identifying conserved T-cell epitopes and broadly neutralizing epitopes to enable pan-ebolavirus vaccines.
[1] Richards PT, et al. Ebola virus' hidden target: virus transmission to and infection of skin. Journal of Virology. 2025. doi:10.1128/jvi.01300-25
[2] Yamaoka S, et al. Ebola virus matrix protein VP40 triggers inflammatory responses linked to the ebolavirus virulence. Proceedings of the National Academy of Sciences of the United States of America. 2026. doi:10.1073/pnas.2508194123
[3] Falasca L, et al. Molecular mechanisms of Ebola virus pathogenesis: focus on cell death. Cell Death and Differentiation. 2015. doi:10.1038/cdd.2015.67
[4] Doghish AS, et al. Host-virus miRNA crosstalk in ebola virus disease. Folia Microbiologica. 2026. doi:10.1007/s12223-026-01538-7
Proteins
Antibodies
ELISA Kits