Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-RA008968A2481phHU |
| Size | US$210 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
| Application | Recommended Dilution |
|---|---|
| WB | 1:500-1:5000 |
| IF | 1:20-1:200 |
Mechanistic target of rapamycin (mTOR) serves as a central integrator of cellular growth signals, coordinating responses to nutrients, energy status, and growth factors. Phosphorylation at serine 2481 represents an autophosphorylation site associated with mTOR catalytic activity, making it a valuable marker for monitoring mTOR complex activation in studies of metabolism, autophagy, and cancer biology.
This recombinant monoclonal antibody, clone 3H11, offers the reproducibility essential for phospho-specific detection work. Generated against a synthetic phosphopeptide corresponding to the human phospho-mTOR (S2481) region, the antibody provides sequence-defined specificity that remains consistent across experiments and over time. This lot-to-lot reliability is particularly important when tracking subtle changes in phosphorylation status across treatment conditions or longitudinal studies.
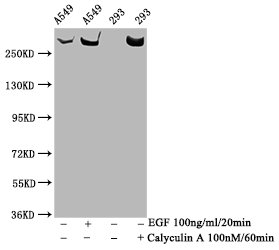
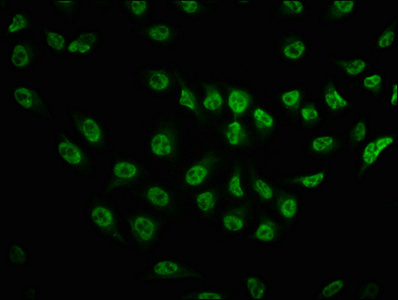
Validation in western blot applications demonstrates robust detection of the expected 289 kDa band in A549 whole cell lysates and HEK293 cells treated with Calyculin A or EGF—conditions known to enhance phosphorylation signals through phosphatase inhibition and growth factor stimulation, respectively. The precise correlation between predicted and observed molecular weight confirms target specificity. For immunofluorescence applications, testing in HeLa cells reveals clear cytoplasmic and perinuclear staining patterns consistent with mTOR's known subcellular localization, with DAPI counterstaining enabling straightforward visualization of signal distribution relative to nuclear compartments.
The antibody's compatibility with both biochemical and imaging approaches provides flexibility for researchers investigating mTOR pathway dynamics in cell biology contexts, from nutrient sensing mechanisms to therapeutic targeting studies in oncology and metabolic disease research.
There are currently no reviews for this product.

